5.4 T细胞转录的可塑性
小鼠中的谱系追踪系统表明,许多亚群的内源性极化的CD4+T细胞在其生命周期中发生了表型的改变,而CD8+T细胞的研究已经证明, 单个初始CD8+T细胞能够同时产生效应性和记忆性淋巴细胞。随后的研究表明,单个CD8+T淋巴细胞在产生具有效应功能的子代时其增殖能力、细胞因子及表型标志物KLRG1和IL-7R的表达等方面不同。令人好奇的是,这种异质性在某种程度上存在组织特异性。另外,将人类单细胞克隆子代的表型分析和TCR测序相结合,揭示了单个T细胞产生的T细胞应答类型具有高度异质性。所有这些研究表明,在单个T细胞生命周期中,它的表型是不稳定的。事实上,单个T细胞具有同时或在不同时间呈现多个T细胞亚群特征的能力,这就产生了T细胞可塑性的概念。重要的是,这种从单个T细胞中产生与不同T细胞亚群相关的不同功能性T细胞的能力似乎有利于宿主免疫。
T细胞免疫表型可塑性的普遍存在很好地体现在Treg细胞上,Treg细胞在应对不同环境时,可以像其他炎性T细胞亚群一样极化。其他辅助T细胞亚群也和Tregs 一样具有可塑性,支持了 T 细胞(炎症和调节)具有较高的灵活性的观点,使其功能可以适应不断变化的环境。与CD4+T细胞相似,CD8+T细胞也具有一定的谱系可塑性。Tc1和CD8+Treg细胞具有相对稳定的特性,而Tc2、Tc9和Tc17细胞则易于获得其他亚群的特性而具有较高的可塑性。Tc2和Tc17细胞的细胞因子谱不变,但能够获得Tc1细胞的额外特性。Tc9细胞在体内相对较不稳定,具有向Tc1或Tc1/Tc2转变的可塑性。
5.4.1 细胞因子—可塑性的主要调节因子
细胞因子在驱动CD4+T细胞亚群之间的可塑性方面发挥着主导作用。TH1细胞与IL-4共同培养或在体内蠕虫感染的情况下使细胞重新极化,产生IL-4并抑制IFNγ的表达。TH2细胞在与IL-12、IFNγ和I型IFNs共同孵育时表达TH1型细胞因子。TH9细胞的发现进一步说明了CD4+T细胞的可塑性。Veldohen等人表明,TGF-β能够重新调节TH2细胞分化为产生IL-9的T细胞而Dardalhon等人证明IL-4能够阻断FOXP3在Treg中的诱导表达,从而诱导产生主要产生IL-9的辅助性T细胞。重要的是,IRF家族的成员能够调节TH9细胞的可塑性。事实上,我们发现IL-1β除了增加TH9细胞的IL-9分泌外,还提高了分化中的TH9细胞的IL-21分泌水平。其潜在机制涉及到STAT1依赖的IRF1激活,并进一步说明了TH9程序的可塑性。
实验表明,Il-1β诱导的TH9细胞在体内维持其转录程序,这提示Il-1β诱导的IRF1表达有助于稳定TH9细胞的转录程序。Th17细胞亚群获得干扰素γ表达或通过失去IL-17和RoR-γt的表达而完全转化为Th1细胞的能力依赖于细胞因子IL-12或IL-23,这两种细胞因子都能激活STAT4。在IL-12、IL-4、IL-6和TGF-β的作用下,极化的小鼠TFH细胞,可诱导产生TH1-、TH2-或TH17型细胞因子,而TH1、TH2、TH17细胞在用IL-21和IL-6处理的TFH细胞环境下培养可表达IL-21、CXCR5以及程序性细胞死亡1 (PD-1)。最后,细胞因子信号抑制基因(SOCS)的特定缺陷可以模拟这些重编程效应,SOCS通过各种机制对抗特定STATs的活性,进一步突出了细胞因子驱动可塑性的重要性。重要的是,细胞因子环境甚至可以影响炎症和调节程序之间的可塑性。例如,TGF-β通过促进FOXP3或IL-10的表达,在TH17细胞向调节表型转化中起关键作用 (Fig5.3)。
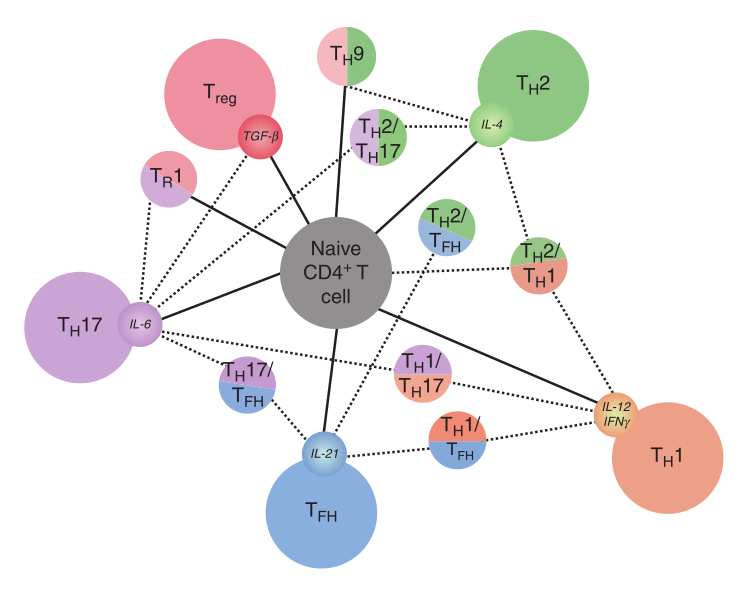
Fig 5.3 细胞因子驱动的T细胞可塑性。关键细胞因子IFNγ、IL-12、IL-4、TGF-β、IL-6和IL-21单独或协同驱动初始CD4+T细胞分化。这些细胞因子能够促进亚群之间的极化或可塑性。
5.4.2 T细胞可塑性的转录和表观遗传调控
继细胞因子在调节T细胞可塑性方面的重要性之后,STATs通过直接响应细胞因子与受体的结合来驱动TH细胞的可塑性。此外,观察发现T细胞主调控因子在转录程序的形成中有重要作用,对实现可塑性也至关重要。然而,对于T辅助细胞亚群极化或可塑性来说,仅有特异性STATs和主转录因子的表达是不够的。其他转录因子,如核受体4A (NR4A)家族、转录调节蛋白BACH2、RUNX蛋白、视黄酸受体和芳基烃受体(AHR),在极化状态的维持中也发挥重要作用,这些转录因子的破坏会导致亚群之间的可塑性增强。染色质的表观遗传调节也会影响T细胞的可塑性。例如,在缺乏HATs的情况下,Tregs细胞失去FOXP3的表达,而获得IL-17的表达。重要的是,一旦效应T细胞的极化功能建立起来,甲基化的减弱将允许T细胞在记忆静止期重新受到刺激后具有显著的可塑性。
像HDACs和H3K27me3相关的抑制复合物1 (PRC1)产生的异染色质,在阻止极性亚群之间的表型可塑性方面也起着关键作用。例如,PRC1对于TH2细胞的功能和限制其向TH1细胞表型的可塑性方面至关重要。对不同极性T细胞亚群中转录可达位点(H3K4me3)或受抑制位点(H3K27me3)相关组蛋白修饰的整体分析,可以为T细胞如何既获得特定功能又保持可塑性提供答案。然而不同T细胞亚群的细胞因子位点显示H3K4me3或H3K27me3标记,染色质结构最极化的转录因子位点包含这两种标记,表明染色质处于允许状态,允许来自相反亚群的转录因子的诱导启动细胞重编程。因此,染色质和DNA修饰酶在T细胞可塑性中具有重要作用,通常是通过与各亚群中的极化转录因子或lncRNAs合作来发挥作用。最后,T细胞的可塑性也可以通过microRNA介导的多种基因的转录后调控产生,如参与细胞因子信号、TCR和共刺激信号以及转录调节通路的基因。
5.4.3 可塑性的代谢调节
T细胞的可塑性可以由细胞的代谢程序控制,该程序可对环境中营养、氧气水平和能量源的波动作出动态响应。免疫应答过程中,当T细胞在不同的微环境(如淋巴器官和组织部位或肿瘤微环境)之间迁移时,这些因素可能很重要。磷脂酰肌醇3激酶(PI3K) -AKT-mTOR通路在环境信号调节可塑性中起重要作用。例如,磷酸酶和张力蛋白同源物的活性使Treg细胞的AKT功能减弱。PTEN的缺失会严重损害Treg细胞的稳定性并促使它们向TH1和TH17细胞转化。此外,尽管已证实Treg细胞中需要一些mTORC1活性,但雷帕霉素治疗实际上促进了Treg细胞的稳定性,Treg细胞中mTORC1的过度激活会导致IL-17的产生和FOXP3表达的缺失。有趣的是,谷氨酰胺是一种重要的生物合成前体,以mTORC1依赖的方式打破TH1和Treg细胞极化之间的平衡。最后,Treg细胞中HIF1α活性可导致异位IFNγ产生和FOXP3表达降低。
5.5 T细胞分化缺陷与人类疾病
如上所述,分化的T细胞亚群对于维持宿主完整性和抵抗感染至关重要。这一概念在遗传缺陷导致T细胞分化缺陷的人类中得到了进一步验证。活化的T细胞表达CD40L,与巨噬细胞和树突状细胞上的CD40结合,导致Th1转变和IL-12的分泌。编码CD40配体的基因突变不仅导致高IgM综合征的发生,其中B细胞分泌除IgM外的所有抗体亚型的能力严重受损,而且还损害了Th1对病原体感染的免疫力。与这些观察结果一致,编码IKKγ(在CD40结合后调节NF-κB活化)基因突变的患者出现单核细胞IL-12分泌受损,从而减少T细胞的IFNγ分泌。重要的是,这些患者对分枝杆菌疾病具有易感性,从而在Th1细胞分化诱导缺陷与疾病易感性之间建立了直接的分子联系。
另一个与T细胞分化缺陷相关的疾病是高IgE综合征(HIES)。这种疾病表现为细菌引起的反复皮肤和肺部感染、真菌引起的念珠菌病以及血清中高浓度的IgE。HIES的发病机制依赖于STAT3基因的突变。如上所述,可由IL-6、IL-22和IL-23诱导表达的STAT3转录因子是Th17细胞分化的核心。Th17细胞极化缺陷导致IL-17A和IL-22分泌减少,这有利于中性粒细胞募集和上皮细胞微生物肽的分泌。IgE水平的增强可能是由于缺乏功能性Th17细胞时Th2反应增强所致。总之,HIES阐明了Th17细胞极化缺陷如何导致疾病发展。这些观察结果与早期发现一致,表明IL-17A和F信号在慢性皮肤粘膜念珠菌病(CMCD)中起着关键作用,该病由白色念珠菌引起,表现为皮肤、指甲、口腔和生殖器粘膜的反复或持续感染。STAT1的功能获得性突变导致Th17细胞极化减少,从而对STAT3激活的细胞因子作出反应,这也与CMCD有关。总的来说,这两种疾病说明CD4 T细胞极化的失调在临床情况下具有高度相关性。
Th2细胞极化与哮喘发生的相关性已在一些临床前研究中得到证实。Th2驱动的IL-4分泌确实促进B细胞类转化为IgE型,这促进了哮喘的发展。这些发现与临床情况相关。在这方面,已经确定了与Th2细胞生物学直接相关的哮喘候选易感基因,包括编码高亲和力IgE受体FcεR1、IL-3、IL-4、IL-5、IL-9、IL-13和GM-CSF的β亚单位的基因。有趣的是,所有后来的细胞因子都能增强IgE类转换、嗜酸性粒细胞存活和肥大细胞增殖,从而支持IgE驱动的过敏反应的发展和维持。因此,编码IL-4基因启动子区域的遗传变异会促进遗传过敏性个体IgE水平的提高。此外,IL-4受体的功能变异与哮喘有关。这些发现同时证明, Th2细胞极化增强可促进哮喘的发展或加重。
调节性T细胞是维持耐受的核心细胞。因此,Treg细胞分化的缺陷在生物体内具有深远的影响。如前所述,转录因子Foxp3是Treg细胞生物学的核心。因此,研究表明scurfy突变(导致Foxp3的DNA结合域丢失)的小鼠在出生后数周内死于大量淋巴组织增生和自身免疫反应,而患有IPEX(免疫失调、多发性内分泌腺病、肠病、X连锁)综合征的患者临床表现与小鼠实验结果基本一致。这种致命的自身免疫性疾病是由FOXP3基因突变引起的。有趣的是,与健康人相比,IPEX患者的循环Treg数并未减少。然而,它们的抑制能力受到损害,这解释了自身免疫的产生。
效应CD8+T细胞功能的缺陷也可能导致疾病的发生。例如,噬血细胞性淋巴组织细胞增多症(HLH)是一种危及生命的疾病,由CD8 T细胞和NK细胞的细胞毒性颗粒释放缺陷引起。然而,这两种细胞类型的细胞因子分泌在HLH患者中得以保留,并且由于促炎症细胞因子的释放增加,加重病情。HLH的变体也被描述为家族性噬血性细胞淋巴组织细胞增生症(FHL)。这些变体都影响CD8 T细胞溶解途径中的特定蛋白质。在这方面,细胞溶解颗粒穿孔素的遗传缺陷与FHL2有关,穿孔素在靶细胞中形成一个孔,从而使其能够被杀死。同样,含有细胞毒颗粒和质膜的囊泡融合所需的蛋白Syntaxin 11和Munc18-2的缺陷分别与FHL4和FHL5有关。CD8 T细胞细胞毒性活性的缺陷也表现在Chediak-Higashi综合征(CHS)中,这是由编码LYST蛋白的基因突变引起的。CHS患者的CTL具有不能降解的细胞毒性颗粒,异常颗粒积聚在细胞溶解活性缺陷的T淋巴细胞中。受影响的个体遭受严重的细菌和真菌感染。总的来说,CD8 T细胞毒性缺陷与人类的几种疾病直接相关。
5.6 结语
T细胞分化成各种具有特殊功能的效应和调节亚群。某些T细胞亚群的高度可塑性不仅可以保护宿主完整性免受多种感染,而且在特定疾病过程中也很重要。例如在沙门氏菌的感染中,它最初定植于肠上皮,从而引发细菌鞭毛蛋白特异性的Th17依赖性反应,导致抑制细菌生长的抗菌蛋白的IL-22依赖性释放。鞭毛蛋白特异性Th1细胞也在感染的这一阶段被检测到,可能来自ex-Th17细胞。细菌随后下调鞭毛蛋白的表达,以逃避IFNγ激活的巨噬细胞的杀伤,并进行系统性传播。然而,在感染的系统性阶段,T细胞对细菌抗原的反应发生了适应性变化,使其能够在巨噬菌体中存活,从而导致Th1细胞极化从而清除细菌。这种免疫反应的时间和环境依赖性适应强调了理解免疫效应子和调节性T细胞亚群分化的关键过程的重要性。在这方面,对分化T细胞亚群的功能基因组分析和单细胞分析能够鉴别可用于治疗目的的新药物靶点。
— THE END —
▉ 往期文章目录
基础免疫学系列
肿瘤免疫学系列
肿瘤免疫学-第3章:CD8+ T细胞在免疫治疗、放疗和化疗中的作用(下)
肿瘤免疫学-第3章:CD8+ T细胞在免疫治疗、放疗和化疗中的作用(上)
生信入门学习笔记系列
基础实验技术系列
Annexin V/PI染色检测细胞凋亡-OncoLab实验室
仪器操作系列
蔡司LSM800激光共聚焦显微镜操作视频-中西医结合基础研究中心
IN Cell Analyzer2200高内涵细胞成像分析系统操作教程-中西医结合基础研究中心
文献分享系列
年度巨献:神刊CA发布最新癌症数据:全球1000万人死于癌症,中国占比超3成
HLA-I分子加工和递呈抗原功能受损是免疫检查点抑制剂耐药的重要原因
作图投稿选刊系列
GraphPad+AI制作可发表级别的柱状图-OncoLab实验室
PubMed中如何按影响因子筛选检索结果并显示影响因子及分区?
如何使用EndNote软件插入参考文献(附6813种杂志参考文献格式文件)
中医中药系列

关注本号~

加入读者交流群~
本篇文章来源于微信公众号: OncoLab
 微信扫一扫打赏
微信扫一扫打赏
 支付宝扫一扫打赏
支付宝扫一扫打赏