7.4 癌症中嘌呤信号转导的临床前证据
7.4.1 肿瘤细胞外ATP
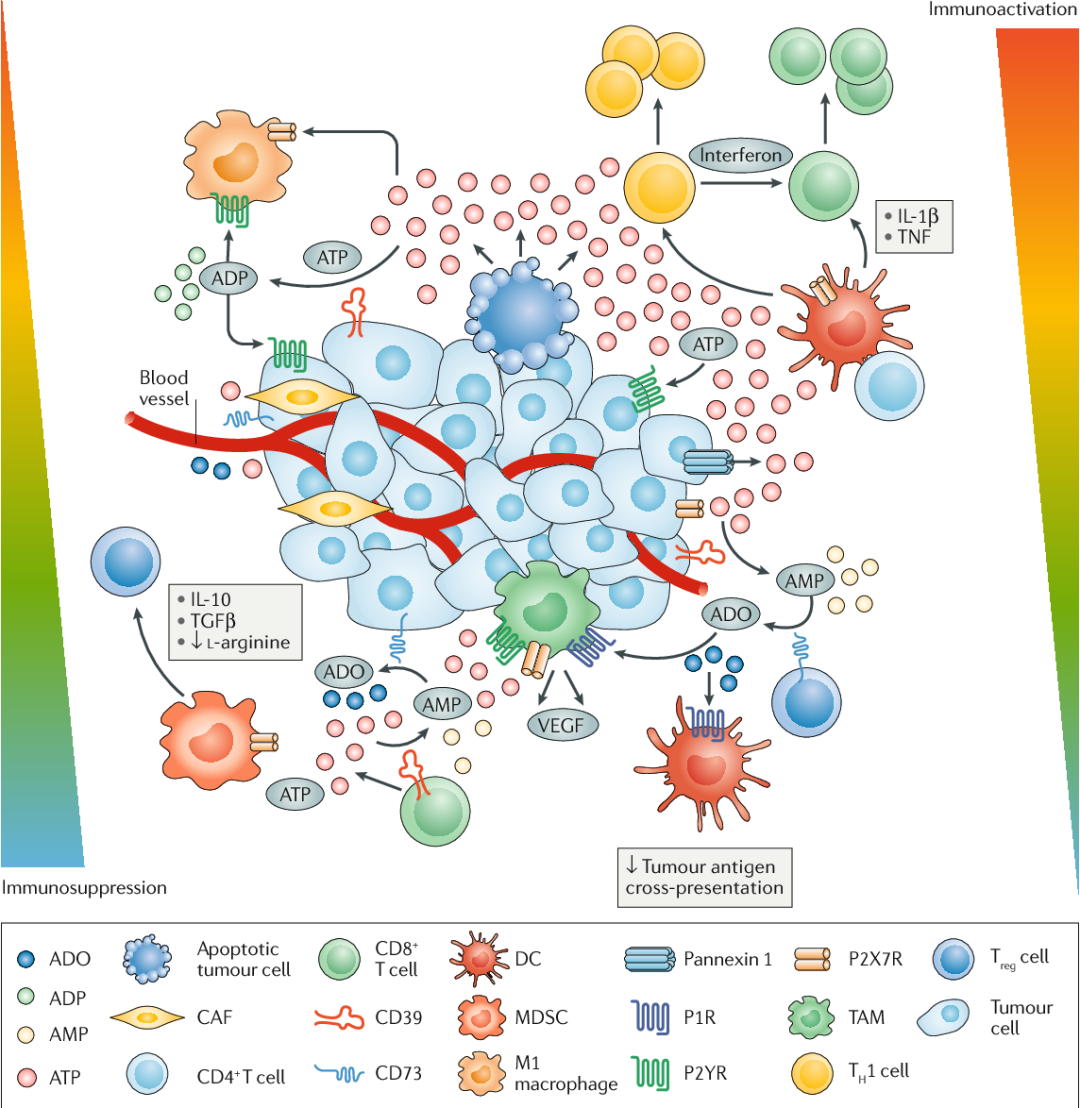
由于TME内细胞外ATP的浓度极高,再加上肿瘤和免疫细胞上P2受体表达的增加,因此嘌呤能途径在直接调节肿瘤生长和抗肿瘤免疫反应中发挥着重要作用。
几项研究表明,P2X7R直接作用于肿瘤细胞可促进其生长,而与宿主细胞(如DC、巨噬细胞和T细胞)的结合则可以激活强烈的抗肿瘤反应。P2X7R的促肿瘤作用包括刺激肿瘤细胞生长,促进MDSC释放免疫抑制因子,以及刺激包括VEGF在内的因子产生介导肿瘤新血管的形成。
因此,前列腺癌细胞中P2X7R沉默可下调参与上皮细胞/间质转化的基因,包括Snail、E-钙粘蛋白、Claudin-1、IL-8和基质金属蛋白酶(MMP-3)。同样,接种小鼠黑色素瘤或人类神经母细胞瘤细胞并敲除P2X7R的小鼠肿瘤生长较慢,转移灶显著减少。与未经治疗的对照组相比,使用选择性抑制剂AZ10606120来抑制P2X7R会显著抑制接种B16F10的小鼠肿瘤生长并减少VEGF的生成。
与上述研究相反,DC细胞上P2X7R的激活可促进抗肿瘤活性。P2X7R 长期以来因其在激活 NLRP3 炎性体和 IL-1β 释放中的作用而闻名。
Virgilio的研究小组证明,与野生型同窝小鼠相比,P2X7R基因敲除小鼠的B16F10黑色素瘤生长和肺转移的速度比较快。产生该效应的部分原因是肿瘤内低水平的IL-1β会导致抗肿瘤免疫细胞募集和浸润到肿瘤中的效率减慢。
相关研究证实,在TME中通过P2X7R激活DC细胞中的NLRP3炎症小体,可对其抗原呈递能力产生积极影响,并增强小鼠的抗肿瘤免疫力。此外,通过炎症小体产生的IL-1β可与IL-23共同促进γδT细胞释放IL-17。这进一步促进了CD8+T细胞释放IFN-γ。重要的是,TP与P2X7R或P2Y2R的结合触发了免疫原性信号,使濒死的癌细胞还能够发挥有效的抗癌疫苗效应。
Zitvogel和Kroemer证明了化疗药物米托蒽醌或奥沙利铂等化疗药物可以诱导死亡的肿瘤细胞释放ATP。这导致DC和T细胞的浸润性增强,从而促进更有效的抗肿瘤免疫。在缺乏P2X7R的小鼠中,大量的免疫原性细胞死亡(ICD)在缺乏 P2X7R 的小鼠中被消除,并且这些小鼠没有特异性CD8+T细胞反应。
相关临床观察证实,P2X7R功能缺失多态性对接受蒽环类临床化疗的乳腺癌患者的预后会产生负面影响。
与P2X7R类似,P2Y2R具有直接的促肿瘤作用。激活MDA-MB-231乳腺癌细胞的P2Y2R可增加其增殖、粘附到内皮细胞的能力以及MMP-9和VEGF的生成。进一步研究表明,P2Y2R基因的敲除抑制了前列腺癌细胞的侵袭和迁移。
具有间变性淋巴瘤激酶(ALK)突变的NSCLC患者通常使用ALK抑制剂进行治疗,如克唑替尼。这些患者的总体应答率为74%,无进展生存期的中位数为10.09个月,最初治疗是有效果的,但后期大多数患者对克唑替尼产生了耐药性。耐克唑替尼的非小细胞肺癌细胞系H3122显示P2YR亚型(P2Y1R、P2Y2R和P2Y6R)的表达增加。这些 P2YR 在激活后导致蛋白激酶 C (PKC) 活性和内皮生长因子受体 (EGFR) 的产生增加。
总体而言,这些发现表明P2YR抑制剂可能是治疗对ALK抑制剂产生耐药性的ALK依赖性NSCLC患者的潜在抗癌药物。
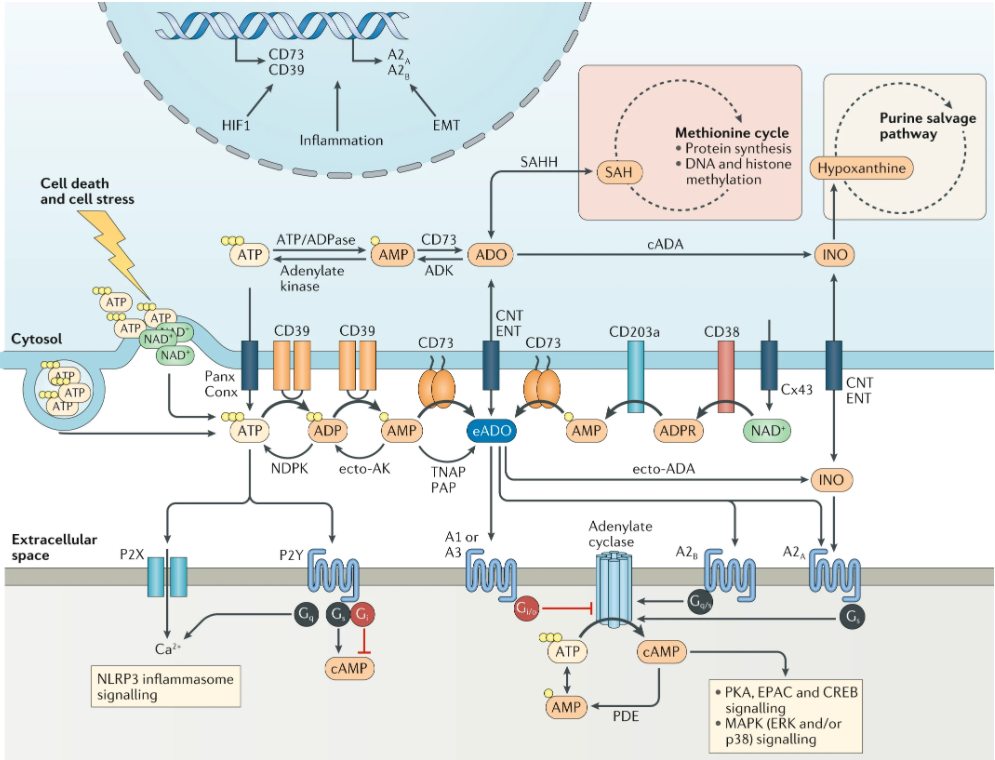
7.4.1.1产生腺苷的外核苷酸酶在癌症进展中的作用
Feng等人使用缺乏CD39的小鼠证明了CD39在促进肿瘤生长中的作用。在该研究中,与相应的WT同龄小鼠相比,CD39基因敲除小鼠的B16F10黑色素瘤生长和转移显著减少。此外,CD39基因敲除小鼠的内皮细胞受损,无法影响肿瘤细胞增殖。其他国家也报告了类似的观察结果。
Sun等人在研究中发现,宿主CD39的缺失消除了B16F10黑色素瘤和MC38结肠肿瘤的实验性肝转移。通过过继性T细胞转移实验表明,Tregs上的CD39表达是肿瘤排斥反应的关键决定因素。将野生型供体的Treg转移到宿主CD39-KO小鼠体内可抑制NK细胞的细胞毒性功能。这项研究也证明了靶向CD39活性可以抑制肿瘤生长和转移。
事实上,CD39的药理学抑制剂POM1可阻止B16黑色素瘤和MC38结肠癌的生长。POM-1的抗肿瘤作用与CD39基因敲除小鼠的抗肿瘤作用相似。但由于POM-1在血液循环中的半衰期较短,所以一些研究小组现已开发出CD39阻断单克隆抗体(MAB)。人类抗CD39单克隆抗体(克隆9-8B)在抑制CD39的酶功能和提高致命转移性患者来源的肉瘤异种移植模型中的存活率方面显示出良好的活性。
类似地,Bastid及其同事描述了另一种抗人CD39单抗(克隆BY40),该单抗在SK-MEL5人黑色素瘤细胞中可抑制CD39酶活性。将该单克隆抗体添加到SK-MEL5和T细胞共培养物中可有效阻断CD39活性并增加CD4+和CD8+T细胞的增殖。然而,这种抗CD39单克隆抗体的治疗效果仍有待于体内试验。
与野生型或杂合子对照组相比,在过表达CD39的患有MC-26结肠癌的小鼠中,转移瘤明显更大。因此,CD39阻断抗体在减轻免疫抑制和恢复对癌症的免疫力方面可能有作用。相反,由于ATP的高积累和雷帕霉素(mTOR)信号激活的机制靶点,老年CD39缺陷小鼠会自发发展为肝细胞癌。
与CD39类似,CD73可以通过多种宿主和肿瘤介导的途径来促进肿瘤生长。Stagg等人使用CD73基因敲除小鼠证明,宿主CD73的缺失赋予了对 B16F10 原发性皮下生长和实验性肺转移的抗性。其他肿瘤模型包括MC38结肠癌、EG7淋巴瘤、AT-3乳腺癌、EL4淋巴瘤和ID8卵巢肿瘤,在缺乏CD73的情况下也可以观察到这种保护作用。宿主CD73缺失对肿瘤影响的大小取决于CD8+T细胞的浸润和活化的程度。
此外,2011年两项独立研究观察得出,与野生型小鼠相比,CD73基因敲除小鼠的Treg抑制效应T细胞产生IFN-γ的作用减弱。在转基因腺癌小鼠前列腺(TRAMP)模型中,宿主CD73的缺失可以保护这些小鼠免于新发前列腺癌;这种保护作用需要IFN-γ、NK细胞和CD8+T细胞。
有趣的是,EG7淋巴瘤模型(相对于亲代EL4淋巴瘤)和B16F10-SIY(相对于亲代B16F10)接种到CD73缺陷小鼠时,会增强对原发性肿瘤的保护作用,这表明肿瘤免疫原性在介导最佳抗肿瘤效应中发挥了作用。
相比之下,CD73缺陷小鼠的抗转移作用由造血细胞或内皮细胞介导。用选择性抑制剂APCP或抗CD73单克隆抗体(克隆TY/23)对CD73进行体内药理学阻断可显著减少肺转移。Terp等人证明,抗人CD73单克隆抗体(cloneAD2)通过诱导表面CD73的内化,以减少人类乳腺癌MDA-MB-231和LM3转移的数量。重要的是,抗转移作用与 CD73 催化功能无关。
此外,目前研究表明,抗CD73单克隆抗体对转移的最佳保护很可能取决于宿主对Fc受体的激活以及肿瘤上CD73的表达水平。
7.4.2 腺苷受体在肿瘤进展和转移中的作用
A1R对肿瘤细胞的影响主要在体外进行研究,在体内研究该受体在肿瘤进展和转移中的作用的数据有限。Synowitz等人的一项研究表明,腺苷通过A1R作用于小胶质细胞会损害GL261胶质母细胞瘤的生长。用A1R激动剂N6环戊基腺苷治疗小鼠可显着减小肿瘤大小,并降低 MMP-2 水平。鉴于MMP-2与促进胶质母细胞瘤的侵袭性相关,本研究得出结论,小胶质细胞上的A1R信号在抑制胶质母细胞瘤发展中至关重要。
A2AR的拮抗剂应用于抗肿瘤治疗的研究备受关注,Ohta等人在大部分A2AR基因靶向小鼠中观察到CL8-1黑色素瘤和RMA T细胞淋巴瘤细胞系的完全排斥反应。本研究使用合成(ZM241385)和天然(1,3,7-三甲基沙芬(咖啡因))A2AR拮抗剂,发现A2AR缺陷小鼠的抗肿瘤功效归因于T 细胞产生的 IFN-γ 增加,以及抑制这些小鼠的血管生成,从而共同促进肿瘤细胞凋亡。在A2AR缺陷小鼠生长的EL4淋巴瘤的排斥反应中,也观察到了类似的结果。
最近的一项研究还表明,A2AR缺陷小鼠可以依赖CD8+T细胞以增强对原发性皮下肿瘤SM1WT1黑色素瘤和AT-3乳腺肿瘤生长的抑制作用。与WT对照组相比,缺乏A2AR的宿主对于介导LWT1黑色素瘤肺转移的保护有重要的作用。
此外,缺乏A2AR的宿主改善了T细胞的记忆反应。在本研究中,皮下接种低剂量的EL4淋巴瘤细胞可以导致A2AR缺陷小鼠和WT小鼠的肿瘤排斥反应,然而,随后用致死剂量的同一肿瘤再次激发仅导致A2AR缺陷小鼠的肿瘤排斥反应,而未导致WT小鼠的肿瘤排斥反应。在过继转移到荷瘤小鼠之前,用A2AR抑制剂ZM-241385培育肿瘤特异性CD8+T细胞可增强其介导肿瘤排斥反应的能力。
siRNA 基因靶向敲除 T 细胞上的 A2AR 可以更好地抑制肺转移并延长 RMA 接种小鼠的存活时间,进而延长接种RMA的小鼠的生存期。许多研究都证明了A2AR抑制剂能有效减少CD73的过度表达和B16F10黑色素瘤的转移,其中NK细胞在限制小鼠肿瘤的转移性生长中起主导作用。
与A2AR类似,Morello的研究小组证明,使用A2BR激动剂BA Y60-6583激活A2BR可促进B16F10黑色素瘤的生长。在这些初步发现的基础上,该研究组进一步证明,激动剂BA Y60-6583刺激A2BR可诱导宿主黑色素瘤相关成纤维细胞表达成纤维细胞相关蛋白(FAP)和成纤维细胞生长因子-2(FGF-2)。这些FAP+成纤维细胞通过释放趋化因子CXCL12触发血管生成,从而增加肿瘤内CD31+内皮细胞的数量。
类似地,使用A2BR抑制剂PSB1115可阻止肿瘤生长,主要是通过减少肿瘤中MDSC的积聚来发挥这些作用。Linden的研究小组表明,使用选择性拮抗剂ATL801阻断A2BR可减缓MB49膀胱癌和4T1乳腺癌的体内生长,这是由于通过IFN-γ-CXCR3轴增加了适应性免疫反应。
同样,与WT对照小鼠相比,A2BR缺陷小鼠在Lewis肺癌小鼠模型中表现出肿瘤生长延迟和生存期延长。这种抑制是由于A2BR缺陷小鼠无法分泌VEGF,从而阻止血管生成。在类似的体外实验中,靶向敲除人A375黑色素瘤细胞上A2BR的siRNA基因可抑制促血管生成细胞因子IL-8的释放。
Mittal等人最近的研究表明,A2BR在肿瘤细胞上的表达,对于促进肺转移至关重要,siRNA 基因靶向敲除 A375 黑色素瘤细胞上的 A2AR 可显著减少这些肿瘤的转移(特别是小鼠三阴性乳腺癌E0771和4T1.2,以及人类三阴性乳腺癌MDA-MB-231)。而宿主A2BR在这些研究中没有显著作用。
据报道,A3R与A2AR和A2BR不同,A3R具有肿瘤抑制功能。这一点在异种移植模型中可以证实,在异种移植模型中,用A3R激动剂LJ-529治疗的小鼠通过诱导细胞凋亡和下调Wnt信号分子Akt、Cyclin D1和GSK-1β来增强抗肿瘤作用。有趣的是,A3R激动剂也能够下调ER+乳腺癌肿瘤(包括MCF7和T47D)上ER-α的表达。
在另一项研究中,当小鼠在化疗前接受A3R激动剂CF101治疗,可观察到中性粒细胞和其他白细胞数量的增加,这表示其增强了人类结肠癌模型中化疗的治疗效果。最后,使用激动剂 CI-IB-MECA 激活 A3R 可抑制 B16F10黑色素瘤生长和血清 IL-12 增加,这是一种促炎细胞因子,对 NK 细胞的活化至关重要。
此外,当来自CI-IB-MECA处理的小鼠的脾细胞被选择性转移时,可观察到转移显著减少,这意味着在该模型中A3R具有抗肿瘤作用 。
7.5 联合疗法
相关临床数据和临床前小鼠肿瘤模型的结果表明,肿瘤中存在多种免疫抑制途径,它们的共同靶向性提高了宿主抗肿瘤免疫的效果。嘌呤能通路在促进肿瘤生长和转移中的作用使其成为肿瘤免疫治疗的新靶点。我们现在讨论一些有关证明该途径的靶向性与化疗或其他免疫检查点分子协同作用的临床前研究。
Allard等人发现,抗CD73单克隆抗体(克隆TY/23)显著增强了抗CTLA-4和抗PD-1单克隆抗体对皮下MC38-OVA(结肠)或RM-1(前列腺)肿瘤小鼠以及携带自发转移性的4T1.2乳腺癌细胞小鼠的活性。这种效应通过激活CD8+T细胞和IFN-γ来介导。
其他两组也报告了类似的发现。Lannone等人报道,与单药治疗相比,用CD73特异性抑制剂、APCP和抗CTLA-4单抗联合治疗显著延缓了B16F10黑色素瘤的生长。Hay等人还表明,用小鼠/人交叉反应性抗CD73单克隆抗体、MEDI9447和抗PD-1治疗可延缓CT26结肠腺癌的生长,从而延长这些小鼠的生存期。
Stagg的研究表明,阻断CD73或A2AR与细胞毒性化疗相结合是有效的。接受抗CD73单抗和阿霉素联合治疗的小鼠在4T1.2乳腺癌中表现出比单药治疗更强的抗肿瘤免疫应答。当一种特殊的A2AR阻断抑制剂SCH58261与阿霉素联合使用时,也能观察到类似的效果。
类似地,可以用A2BR抑制剂PSB1115和化疗药物达卡巴嗪或吉西他滨联合治疗小鼠,其机制是通过减少TME内MDSC的积累,来显著抑制B16F10黑色素瘤的生长。当使用可溶性B7-DC/Fc融合蛋白治疗时,A2AR缺陷小鼠的无瘤生存期和总生存期比WT小鼠长,该融合蛋白特异性地针对树突状细胞上表达的PD-1受体,导致T细胞活化。
Mittal等人报告在患有实验性肺转移的小鼠中,SCH58261和抗PD-1联合治疗可显著降低转移水平。其中Beavis等人最近的一项研究表明,可通过将抗PD-1与A2A缺陷嵌合抗原受体(CAR)-T细胞结合,或用拮抗剂SCH58261来治疗WT CAR T细胞,此疗法可显著提高T细胞对表达HER2的小鼠肿瘤的抗肿瘤功效。
此外,Iannone等人表明,A2AR拮抗剂ZM241365的药理阻断可提高B16F10黑色素瘤模型中抗CTLA-4治疗的疗效。在这项研究中,作者还报告了A3激动剂CI-IB-MECA与抗CTLA-4协同抑制黑色素瘤荷瘤小鼠的肿瘤生长。最后,Young等人表明,联合抑制A2AR和抗CD73单抗可显著降低小鼠的肿瘤生长和肺转移。嘌呤能途径内的共同靶向分子提高了抗转移效力,这表明这些分子在介导抗肿瘤免疫中的非冗余性质。
7.6 正在进行的临床试验
减少肿瘤内微环境缺氧的最简单方法之一是增加氧气水平。实验研究表明,与接受100%氧气或空气的小鼠相比,在照射前和照射期间,允许在空气中呼吸高氧(95%氧气+5%二氧化碳)的荷瘤小鼠对放射治疗的反应显著增强,然而,最近的研究表明,高浓度补充氧气会导致氧中毒和非特异性炎症反应。
Sitkovsky的研究小组表明,向小鼠补充60%的氧气,以诱导呼吸性高氧,可显著减少小鼠的肺MCA205、B16F10和4T1肿瘤转移的数量。且由于呼吸性高氧广泛应用于临床环境,本研究的作者得出结论,用其他可用的免疫疗法补充60%的氧气可能是治疗癌症的一种新策略。
除了缺氧,临床上还有大量的靶向嘌呤能受体的相关研究,这些受体影响细胞外ATP和腺苷的生成。目前,一些研究评估了人类癌症中嘌呤能受体的表达水平,该表达与这些患者的无复发生存率(RFS)和总生存率(OS)有关(Table 7.2)。这些关联研究加上强有力的临床前研究,促进许多有关人类癌症嘌呤能途径的抗肿瘤疗效的临床试验的启动。
例如,使用多克隆羊抗P2X7抗体(BIL010t)的一期临床试验于2014年完成(NCT02587819)。21例基底细胞癌患者经28天局部施用BIL010t后,其中65%的患者病变面积缩小。切除病变的组织病理学显示2例完全缓解,9例部分缓解,8例无反应。
此外,Medimune Inc.最近报道了MEDI9447的开发,这是一种小鼠/人类交叉反应性抗CD73单克隆抗体,用于增强临床前肿瘤模型中的抗肿瘤免疫。目前正在进行使用 MEDI9447与(MEDI473)的一期试验(NCT02503774)。最后, Corvus Pharmaceuticals,Inc.和Palobiofarma已经开发出用于治疗癌症的A2AR抑制剂,即CPI-444 (NCT02655822) 和PBF-509(NCT02403193)。
此外,CPI-444将在晚期肿瘤患者中与阿替唑珠单抗(抗PD-L1)合用,而Palobiofarma的目的是在非小细胞肺癌患者中测试PBF-509与抗PD-1治疗的联合阻断作用。
— THE END —
▉ 往期文章目录
基础免疫学系列
肿瘤免疫学系列
肿瘤免疫学-第6章:调节性T细胞的作用、作用机制以及对癌症的影响(下)
肿瘤免疫学-第6章:调节性T细胞的作用、作用机制以及对癌症的影响(上)
肿瘤免疫学-第3章:CD8+ T细胞在免疫治疗、放疗和化疗中的作用(下)
肿瘤免疫学-第3章:CD8+ T细胞在免疫治疗、放疗和化疗中的作用(上)
文献分享系列
年度巨献:神刊CA发布最新癌症数据:全球1000万人死于癌症,中国占比超3成
HLA-I分子加工和递呈抗原功能受损是免疫检查点抑制剂耐药的重要原因
生信入门学习笔记系列
基础实验技术系列
Annexin V/PI染色检测细胞凋亡-OncoLab实验室
仪器操作系列
蔡司LSM800激光共聚焦显微镜操作视频-中西医结合基础研究中心
IN Cell Analyzer2200高内涵细胞成像分析系统操作教程-中西医结合基础研究中心
作图投稿选刊系列
GraphPad+AI制作可发表级别的柱状图-OncoLab实验室
PubMed中如何按影响因子筛选检索结果并显示影响因子及分区?
如何使用EndNote软件插入参考文献(附6813种杂志参考文献格式文件)
中医中药系列

关注本号~

加入读者交流群~
本篇文章来源于微信公众号: OncoLab
 微信扫一扫打赏
微信扫一扫打赏
 支付宝扫一扫打赏
支付宝扫一扫打赏