人类行为的多样性源于复杂精密的感官系统与大脑的协作。大脑作为一个高度灵活的神经中枢,通过从持续的感觉信号流中筛选出对个体最重要的外部和内部环境变化,来实现感知和应对。大脑不仅能够有效地组织这些感觉信息,用于认知、行动、决策、审美以及记忆存储,还可以有选择性地忽略无关信息。同时,它能通过神经信号将这些复杂的处理结果及其心理表现传递给其他大脑区域,从而实现协调与反馈。
单个神经元是大脑的基本信号单位。人脑中约有860亿个神经元,这些神经元可以分为至少一千种不同类型。然而,决定人类行为复杂性的并不仅仅是这些神经元的多样性,更关键的是它们通过精确的解剖回路组织起来。事实上,大脑的核心组织原则在于:即使神经元的性质相似,不同的连接模式仍然能够产生多样的行为反应。
神经系统的组织原则虽然相对简单,但其功能极为复杂。理解神经系统如何产生行为,主要可以从以下五个方面入手:
1. 单个神经细胞的结构;
2. 神经元内部及彼此之间的信号传递机制;
3. 神经元与其靶标(如肌肉和腺体效应器)之间的连接方式;
4. 不同连接模式如何与特定行为类型相关;
5. 经验如何改变神经元及其连接。
本书的内容将围绕这五个主题展开。本章首先概述行为的神经控制原理,介绍神经元的结构与功能以及支持神经元的胶质细胞。接着,我们将探讨单个神经元如何传递信号,以及多个神经元如何通过连接产生简单行为,如膝跳反射。随后,我们将延伸到更复杂的行为调控,这些行为由复杂且灵活的神经回路来管理和调节。
神经系统主要由两类细胞组成:神经元和神经胶质细胞。
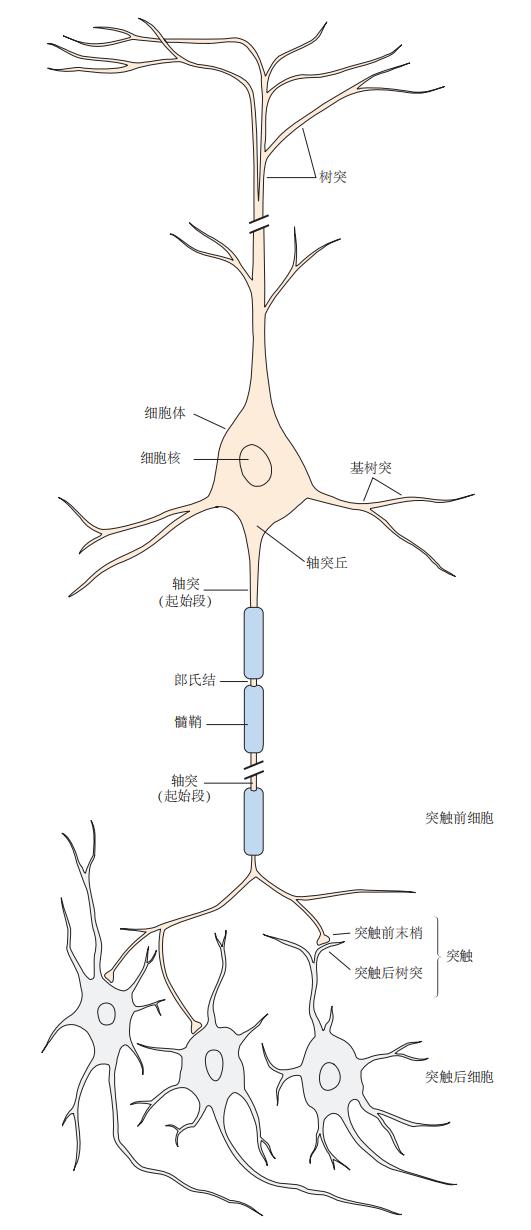
典型的神经元由四个形态学上可区分的区域构成:细胞体、树突、轴突和突触前末梢(图3.1.1)。每个区域在信号的生成和与其他神经元的通信中发挥着独特作用。
细胞体是神经元的代谢中心,包含细胞核,核内存有细胞的遗传物质。细胞体中还有内质网,它负责合成细胞所需的蛋白质。细胞体延伸出两种不同的结构:多个短小的树突和一条较长的轴突。树突呈树枝状分叉,是接收来自其他神经元信号的主要部位。轴突通常在分支之前从细胞体延伸,能够将信号传递给多个目标神经元。轴突可以在0.1毫米到1米的距离内传导电信号。这些电信号,也叫动作电位,从轴突起始段的一处特殊触发区域产生,并以每秒1米至100米的速度沿轴突传播。由于动作电位是一种全有或全无的脉冲信号,它在传播过程中不会衰减或失真,因此电信号的幅度始终保持在约100毫伏,沿轴突周期性地再生(图3.1.2)。
动作电位是大脑接收、处理和传递信息的基本信号。虽然这些信号由不同的环境刺激(如光、触觉、气味、声波等)引发,但它们在神经系统中本质上是相同的。无论是传递视觉信息的信号,还是传递嗅觉信息的信号,它们的生理形式一致。这揭示了大脑功能的一个关键原则:动作电位所传递的信息类型并不取决于信号的形式,而取决于信号传播的通路。大脑通过分析和解读特定神经通路中的信号模式,来生成我们所体验的视觉、听觉、触觉、嗅觉和味觉。
图 3.1.1:神经元的结构
脊椎动物神经系统中的大多数神经元具有一些共同的主要特征。细胞体包含细胞核,是遗传信息的存储中心。细胞体衍生出两种类型的结构:轴突和树突。轴突是神经元的传递元件,其长度变化很大,最长的轴突在体内可延伸超过1米。相比细胞体的直径(约50微米或更大),中枢神经系统中的大多数轴突相对较细,直径在0.2微米至20微米之间。许多轴突外部被一层脂肪质的髓鞘包裹,该髓鞘在郎飞结处定期中断,允许信号的快速传导。动作电位,神经元的信号传递形式,从轴突的起始段发出,并传播至突触,这是信号从一个神经元传递给另一个神经元的地方。突触前神经元的轴突末梢通过突触将信号传递给突触后细胞。一个轴突的分支可与多达一千个突触后神经元形成连接。树突和细胞体一起作为神经元的输入结构,负责接收来自其他神经元的信号。
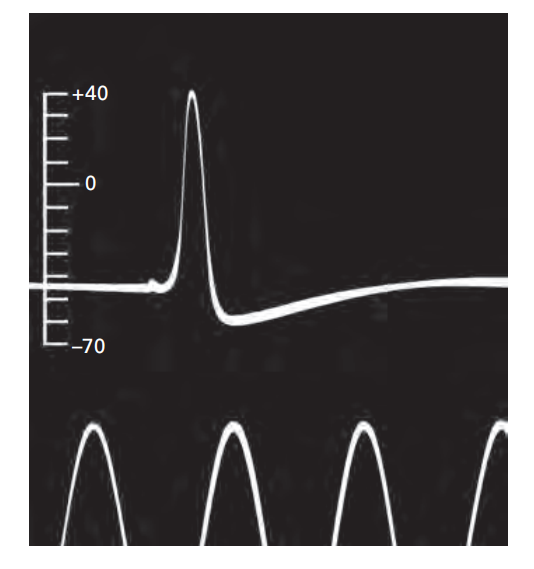
图 3.1.2:动作电位的首次记录
1939年,艾伦·霍奇金和安德鲁·赫胥黎首次使用装满海水的玻璃毛细管电极记录了乌贼巨型轴突的动作电位。图中的定时脉冲(底部)间隔2毫秒。垂直刻度显示细胞内部电位的变化,单位为毫伏,外面的海水作为零电位参考点。
为了加快动作电位的传导速度,较粗的轴突被包裹在一种称为髓磷脂的脂质层中,形成绝缘鞘。这种绝缘鞘在轴突上的定期间断点称为郎飞结,这些点是动作电位得以再生的未绝缘区域。(有关髓鞘形成的详细讨论见第7章和第8章,动作电位的机制详见第10章。)
当轴突接近末端时,它会分裂成更小的分支,并在被称为突触的特殊通信区域与其他神经元接触。传递信号的神经元称为突触前细胞,接收信号的神经元则称为突触后细胞。突触前细胞通过其轴突末端的突触前末梢(或称为神经末梢)来传递信号。突触前细胞和突触后细胞之间通过一个极窄的空间分隔,称为突触间隙。大多数突触前末梢与突触后神经元的树突连接,少部分与细胞体相连,极少数情况下会终止在突触后细胞的轴突起点或末端(见图3.1.1)。一些突触前神经元通过激发来作用于突触后细胞,而另一些则通过抑制来影响其目标细胞。
神经元学说(详见第1章)提出,每个神经元都是一个独立的细胞,具有独特的形态,且是神经系统信号传递的基本单位。回顾这一学说的发展过程,我们能够感受到科学家们最初提出这一基本概念时面临的巨大挑战。与其他可在显微镜下轻易观察到的细胞不同,神经元的形态复杂,尤其是其树突的复杂分支和轴突的漫长延展,使得早期科学家难以厘清它们之间的关系。尽管1830年代雅各布·施莱登和西奥多·施旺提出了细胞理论,认为细胞是所有生命体的基本结构单元,这一理论成为了生物学的基石,但大多数解剖学家依然不接受该理论适用于大脑。他们认为大脑是由连续、网状的纤细结构组成的复杂体系。
直到19世纪后期,神经元的独立性结构才变得清晰。这得益于拉蒙·卡哈尔采用了由高尔基引入的银染法。这种方法至今仍然使用,具有两个显著优点。首先,银染法通过一种尚未完全理解的随机机制,仅染色大脑中约1%的神经元,使研究者能够在不受邻近细胞干扰的情况下观察单个神经元。其次,这种染色方法能够清晰显示神经元的完整形态,包括细胞体、轴突和所有树突分支。银染法的结果显示,神经元之间不存在细胞质的连续性,卡哈尔由此预见性地得出结论,即便是在突触处,神经元之间也没有直接的连续性。
拉蒙-卡哈尔不仅将高尔基染色法应用于人类胚胎的神经系统,还将其推广到多种动物的胚胎神经系统中。他通过观察神经系统各个部位的神经元结构,成功描述了神经细胞的种类,并绘制出许多神经细胞之间的精确连接方式。通过这些研究,除了神经元学说外,拉蒙-卡哈尔还提出了两个在理解神经系统通信方面具有重要意义的原则。
第一个原则是动态极化原理,该原理指出神经细胞内的电信号具有方向性,通常从神经元的突触后部位(主要是树突和细胞体)向轴突的触发区域流动。在触发区域产生动作电位后,信号沿着轴突的整个长度传播,直到轴突末端。在大多数已研究的神经元中,电信号都是沿轴突单向传导的。
卡哈尔提出的第二个原则是连接特异性原理,它指出神经元在形成神经网络时不会随机连接,而是按照特定的方式在特定接触点与特定的突触后细胞建立联系。并非所有细胞都能彼此随意连接。这两个原则,即动态极化和连接特异性,是现代研究大脑神经回路的重要理论基础。
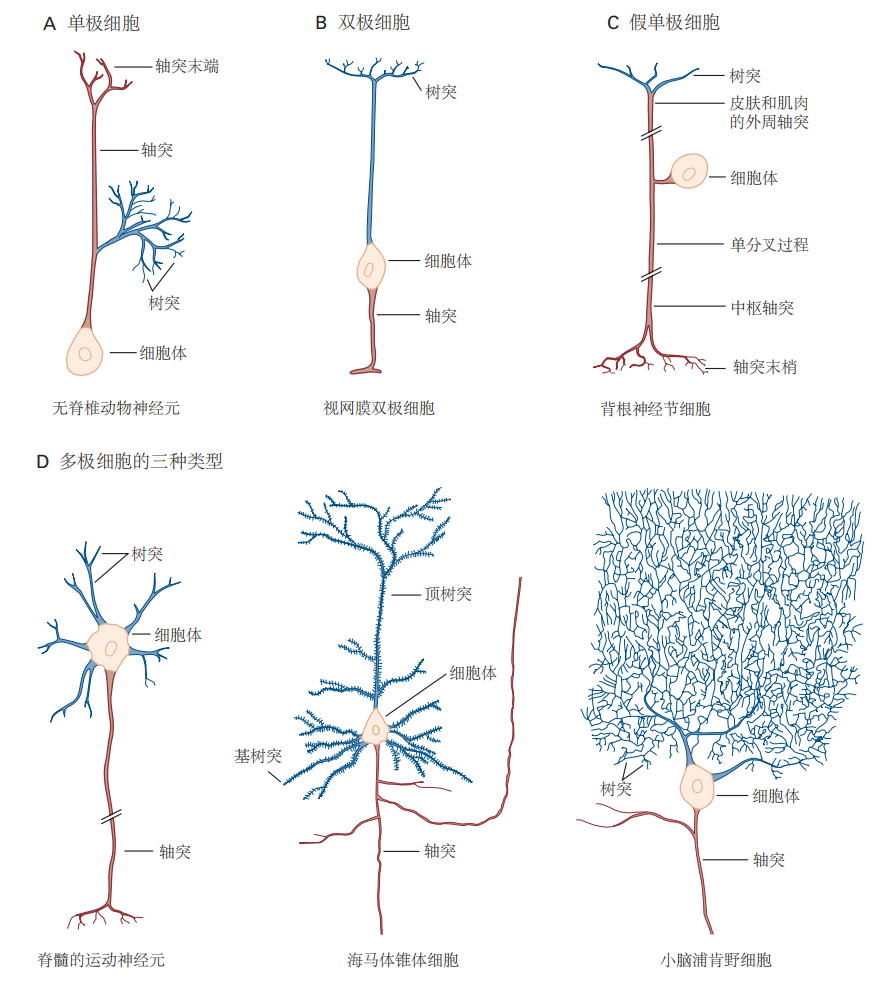
此外,卡哈尔还率先认识到,区分神经元类型的最重要标准是其形态,尤其是细胞体突起的数量。根据这一特征,神经元可以分为三大类:单极神经元、双极神经元和多极神经元。其中,单极神经元是最简单的类型,只有一个主要突起,该突起通常会分支为多个结构,其中一个作为轴突,其他分支用于接收信号(见图3.1.3A)。这种细胞类型在无脊椎动物的神经系统中占主导地位,而在脊椎动物中则主要出现在自主神经系统中。
双极神经元具有一个椭圆形的细胞体,并从中产生两个不同的突起:一个是接收其他神经元信号的树突,另一个是将信息传递至中枢神经系统的轴突(见图3.1.3B)。许多感觉神经元属于双极神经元,例如视网膜的双极细胞和鼻腔的嗅觉上皮细胞。将触觉、压力和疼痛信号传递到脊髓的感受器神经元最初也是双极细胞,但它们的两个突起会在发育过程中融合,形成一个连续的单一结构,从细胞体的一个点发出。此时,原本的树突发生结构上的特化,变得类似于轴突。这类细胞被称为伪单极细胞,它们的一侧轴突负责将信息从皮肤、关节和肌肉中的感觉受体传递到细胞体,另一侧则将这些感觉信息传递至脊髓(见图3.1.3C)。
多极神经元在脊椎动物的神经系统中占主导地位。这类神经元通常有一个轴突和多个从不同位置发出的树突(见图3.1.3D)。多极细胞的形态差异显著,特别是在轴突长度、树突的分支程度及其复杂性方面。一般来说,树突的分支越多,与该神经元形成突触的其他神经元数量也就越多。例如,脊髓运动神经元拥有较少的树突分支,能接受大约1万个突触输入,其中1000个位于细胞体,9000个位于树突上。而小脑的浦肯野细胞具有更加复杂的树突结构,能接收多达100万个突触输入。
根据功能,神经元分为三大类:感觉神经元、运动神经元和中间神经元。感觉神经元负责将外界传感器接收到的信息传递到中枢神经系统,以便进行感知和运动协调。部分初级感觉神经元也被称为传入神经元,这两个术语可互换使用。”传入“指所有从外周传向中枢神经系统的信息,无论这些信息是否产生感知。而”感觉“特指将信息从感觉上皮、关节感觉受体或肌肉传递到中枢神经系统的传入神经元,且这一概念已经扩展,涵盖了大脑皮层初级和次级区域的神经元,它们能够对诸如物体在空间中的移动、声音频率变化或头部角度旋转(通过耳内前庭器官)等感知特征做出反应,甚至包括面部表情等复杂现象。
“传出“一词适用于所有从中枢神经系统传向运动器官的信息,无论这些信息是否导致实际动作。运动神经元将来自大脑或脊髓的命令传递给肌肉和腺体(即传出信息)。传统上,运动神经元被定义为直接激发肌肉活动的神经元,但如今这个定义已扩展,包括那些虽然不直接支配肌肉,但间接指挥动作的神经元。无论是运动神经元还是感觉神经元,它们的共同特征是与外界刺激或身体运动的动态变化保持高度的时间一致性。感觉神经元为大脑提供数据,运动神经元将大脑的指令转化为实际行动。它们共同构成了我们与外部世界的纽带。
中间神经元包含最多的功能类别,并可分为两类:中继中间神经元和局部中间神经元。中继中间神经元(或投射中间神经元)具有较长的轴突,能够在较长距离内将信号从一个大脑区域传递到另一个区域。而局部中间神经元的轴突较短,主要与局部回路中附近的神经元连接。
由于几乎每个神经元都可以被视为中间神经元,这一术语通常用于区分两类神经元:一种是投射到局部回路内的神经元,另一种则是投射到其他神经结构的神经元。有时,该术语也被简化用于指代抑制性神经元,尤其是在研究皮层回路时,但为了明确,应在适当情况下使用“抑制性中间神经元”这一术语。
每个功能分类还可以进一步细分。感觉系统中的中间神经元可以根据它们响应的感觉刺激类型来分类,而这些初步分类还可以依据位置、密度、大小和基因表达模式进一步划分。
事实上,随着信使核糖核酸(mRNA)序列分析技术的进步,我们能够对单个神经元进行分子表型分析,从而更好地了解神经元的复杂性。最新的研究表明,神经元类型的多样性远比之前所认为的要复杂得多(见图3.1.4)。
图 3.1.3:神经元类型的分类
根据从细胞体发出的突起数量,神经元可以分为单极、双极和多极。
A. 单极细胞具有一个从细胞体发出的单一突起,不同部分负责信号接收或传递。这种细胞类型多见于无脊椎动物。
B. 双极细胞有两个特化的突起:树突接收信号,轴突传递信号。
C. 伪单极细胞是双极细胞的变体,负责将体感信息传递至脊髓。发育过程中,双极细胞的两个突起融合,形成单一的突起,分为两个功能不同的部分:一个延伸至外周(皮肤或肌肉),另一个延伸至中枢(脊髓)。
D. 多极细胞具有一个轴突和多个树突,是哺乳动物神经系统中最常见的神经元类型。举例来说,脊髓运动神经元支配骨骼肌纤维;锥体细胞形似三角形,树突从顶端和基底部发出,存在于海马体和大脑皮层中;小脑的浦肯野细胞以其复杂的树突网络著称,能够接收大量的突触输入。
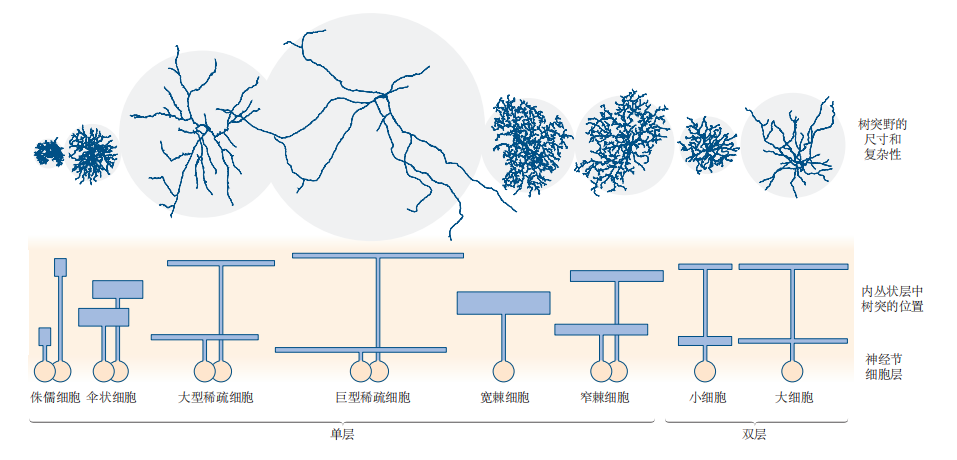
图 3.1.4:感觉神经元的分类
感觉神经元可以根据功能进一步细分。例如,视网膜中至少有13种不同类型的神经节细胞,它们根据树突的大小、形状以及接收输入信号的视网膜层次来区分。内丛状层是视网膜中,双极细胞、无长突细胞和神经节细胞之间的连接区域。
胶质细胞的数量远远超过神经元,在脊椎动物中枢神经系统中,胶质细胞的数量是神经元的2到10倍。尽管其名称源自希腊语中的“胶水”,但实际上,胶质细胞并不只是起到粘合神经细胞的作用。相反,胶质细胞围绕着神经元的细胞体、轴突和树突。
在形态上,胶质细胞不同于神经元,它们没有树突和轴突;在功能上也有明显差异。虽然胶质细胞和神经元来自相同的胚胎前体细胞,但胶质细胞的膜特性不同于神经元,因此不具备电兴奋性,也不直接参与电信号的传递。然而,胶质细胞在帮助电信号沿轴突快速传导方面起着重要作用,同时还在早期发育中指导神经连接以及在学习过程中形成新的或改变的神经连接的稳定性方面扮演关键角色。在过去的十年中,科学界对胶质细胞多功能性的兴趣大大增加,研究已经从认为它们是“支持细胞”转向认识到它们是神经元的功能伙伴(详见第7章)。
3.2 每个神经细胞都是调节特定行为的回路的一部分
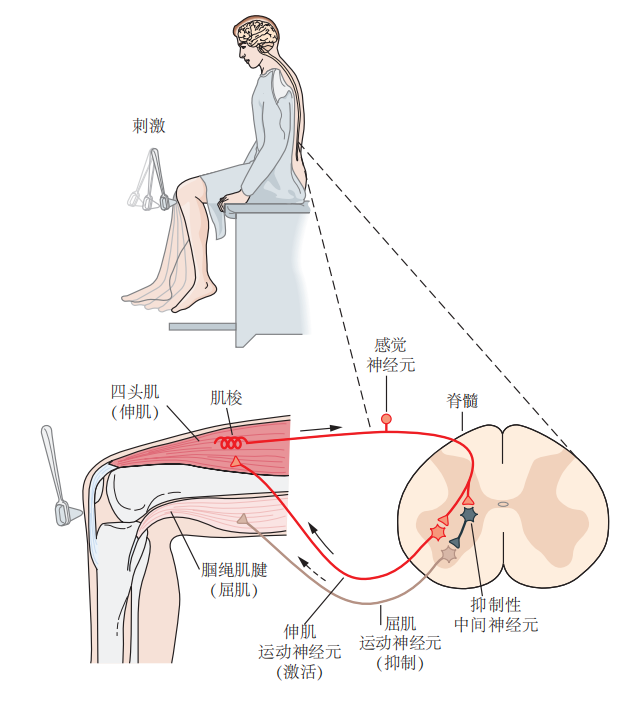
每种行为都由一组特定的相互连接的神经元来调节,每个神经元的功能取决于它与其他神经元的连接。一个典型的例子是膝跳反射。当身体暂时失去平衡时,腿部的股四头肌被拉伸,触发反射。这种拉伸产生的感觉信息传递到运动神经元,运动神经元再向伸肌发出收缩命令,从而帮助恢复平衡。
在临床上,膝跳反射用于检测神经系统的完整性以及脊髓对反射幅度(增益)的控制。这一反射机制十分重要,能够维持股四头肌的正常张力,防止膝盖在站立或行走时屈曲。股四头肌是一块负责伸展小腿的肌肉,其肌腱通过髌骨附着在胫骨上。轻轻敲击髌骨下方的肌腱会导致股四头肌拉伸,触发反射性收缩,产生膝跳动作。通过增加特定肌肉群的张力,牵张反射改变了腿的位置,导致其突然向外伸展(见图3.2.1)。
图 3.2.1:膝跳反射是由感觉神经元和运动神经元组成的一个简单神经回路控制的。当使用叩诊锤轻叩膝盖骨下方时,会拉动股四头肌的肌腱,导致股四头肌——负责伸展小腿的肌肉——被拉伸。此时,肌肉的拉伸信息通过感觉神经元传递至中枢神经系统。在脊髓中,感觉神经元与伸肌的运动神经元形成兴奋性突触,使股四头肌收缩。同时,感觉神经元通过中间神经元抑制拮抗肌(如绳肌)的运动神经元,防止屈肌收缩。这种双重作用产生了膝跳反射的行为。在图中,每个伸肌和屈肌的运动神经元代表了许多神经细胞群。
参与膝跳反射的感觉神经元的细胞体集中在脊髓附近的背根神经节中。这些神经元是伪单极细胞,其一个分支延伸至股四头肌,另一个分支进入脊髓。与股四头肌接触的分支负责感受肌肉的拉伸,当肌肉被拉伸时,肌梭感受器被激活。进入脊髓的分支则与控制股四头肌收缩的运动神经元相连,此外还与抑制屈肌的运动神经元的局部中间神经元连接(见图3.2.1)。尽管这些局部中间神经元并未直接参与反射动作,但它们通过协调对抗肌群的活动,提高了反射动作的稳定性。
因此,产生牵张反射的电信号携带了四种重要信息:
1. 感觉信息从肌肉传递到中枢神经系统(脊髓)。
2. 来自中枢神经系统的运动命令传递至膝跳反射的肌肉。
3. 向拮抗肌的运动神经元发出抑制指令。
4. 与膝跳反射相关的局部神经元活动信息被传递到中枢神经系统的更高层,帮助大脑协调其他行为。
大脑还可以根据情境调节膝跳反射的强度。例如,当我们跑步时,绳肌会弯曲膝盖,拉伸股四头肌。此时,大脑和脊髓会抑制牵张反射,使股四头肌放松。如果这些下行通路被破坏,例如在中风的情况下,反射会过度增强,导致关节僵硬。
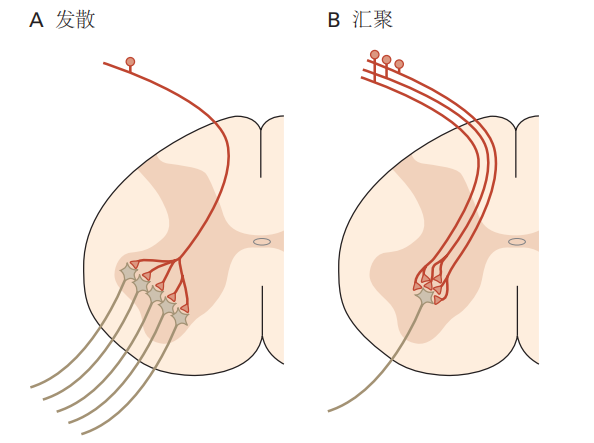
在膝跳反射中,拉伸一块肌肉(股四头肌)会激活数百个感觉神经元,每个感觉神经元与45到50个运动神经元直接连接。此种一个神经元影响多个目标细胞的连接模式称为发散(见图3.2.2A),它在神经系统的输入阶段非常常见。通过将信号分配给多个靶细胞,单个神经元可以产生广泛的影响。相反,单个运动细胞接受来自约130个感觉神经元的200至450个输入联系,这种模式称为收敛(见图3.2.2B)。这种连接模式在神经系统的输出阶段很常见,它能够整合来自多个感觉来源的信息。每个感觉神经元的输入单独较弱,因此收敛有助于确保只有当足够多的感觉神经元一起被激活时,运动神经元才会被触发。
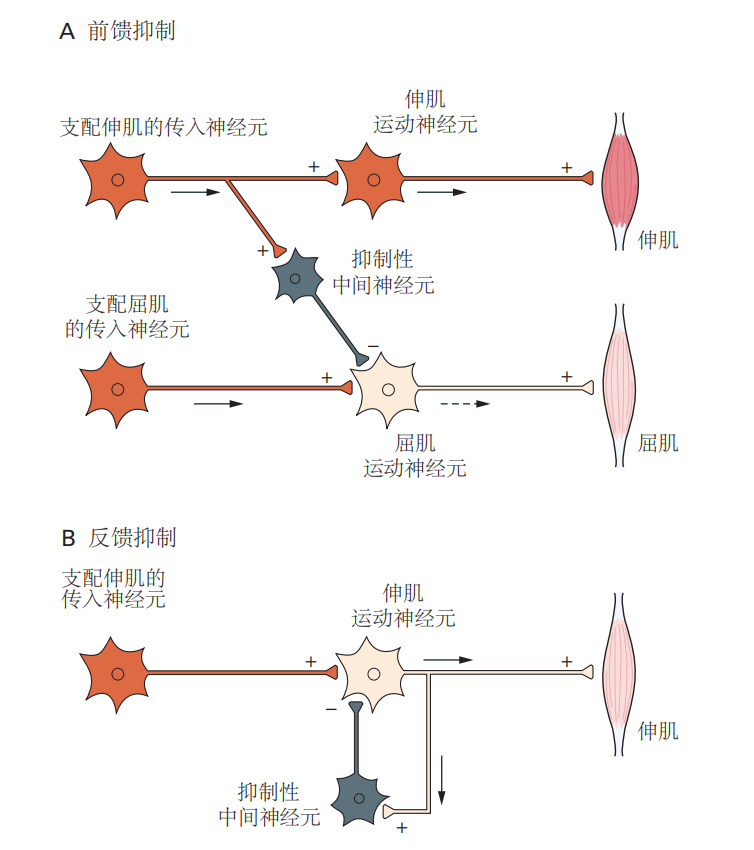
图 3.2.2:牵张反射(如膝跳反射)是一种由两类神经元通过兴奋性突触连接而形成的简单行为。然而,大脑中并非所有信号都是兴奋性的。许多神经元会发出抑制性信号,以减少其他神经元的放电可能性。即使在简单的膝跳反射中,感觉神经元也会形成兴奋性和抑制性的连接。对伸肌的兴奋性连接导致肌肉收缩,而通过抑制性中间神经元对屈肌的抑制性连接则防止了拮抗肌(屈肌)的收缩。这个过程是前馈抑制的一个例子(图3.2.3A)。在膝跳反射中,前馈抑制确保在伸肌收缩的同时,屈肌被抑制,保证了膝关节的协调运动。
一些神经回路则提供反馈抑制。例如,一个运动神经元不仅可以通过兴奋性连接使肌肉收缩,还能通过与抑制性中间神经元的连接,调节自身对肌肉的刺激能力(图3.2.3B)。当运动神经元兴奋时,抑制性中间神经元被激活,进而限制运动神经元的刺激强度。在后面的章节中,我们会遇到更多复杂行为中的前馈和反馈抑制机制。
图 3.2.3:神经元的发散和收敛连接是大脑的一个关键组织特征。
A. 在感觉系统中,通常一个感受器神经元会连接多个代表第二级处理阶段的神经元。随着处理的推进,传入信号会进一步发散,使来自单一感受部位的感觉信息可以广泛分布到脊髓和大脑。
B. 相比之下,运动神经元通常是收敛连接的目标。多个突触前细胞的输入需要汇聚在一起,才能激活运动神经元。这种结构确保了运动神经元的激活需要来自多个来源的信号输入。
本文翻译整理自Principles of Neural Science, Sixth Edition,翻译初稿来自于GitHub/OpenHUTB,本公众号进行进一步校对排版后发布,仅用于学习交流使用,如需购买原版书籍可点击阅读原文购买。
— THE END —
来源 | Principles of Neural Science, Sixth Edition
图文排版 | 李芋岭
审核修改 | 王坤
(转载请保留此部分内容)

关注本号~

加入读者交流群~
(添加请备注单位姓名)

加入知识星球~
点亮赞与在看
让更多人看到
本篇文章来源于微信公众号: OncoLab
 微信扫一扫打赏
微信扫一扫打赏
 支付宝扫一扫打赏
支付宝扫一扫打赏