导 读
本章介绍T细胞的抗原识别分子,即T细胞受体(TCR)。上图概述了TCR组装的主要步骤。如第2章所述,TCRs识别肽抗原有一个必要条件,那就是该抗原需要由自身主要组织相容性复合物(MHC)分子进行展示,没有与MHC结合的抗原是不能被TCR识别的,也就是说,它们具有双重特异性。这与B细胞抗原识别分子(抗体)不同,抗体是直接结合各种肽和非肽抗原。
本章最后讨论了TCR和MHC抗原之间如何通过相互作用形成免疫突触的一部分。尽管抗原识别机制不同,但TCR在结构上与免疫球蛋白相似(Fig 7.1)。像B细胞抗原受体一样,TCR也是呈克隆分布。T细胞的每个克隆都表达不同的抗原受体分子。
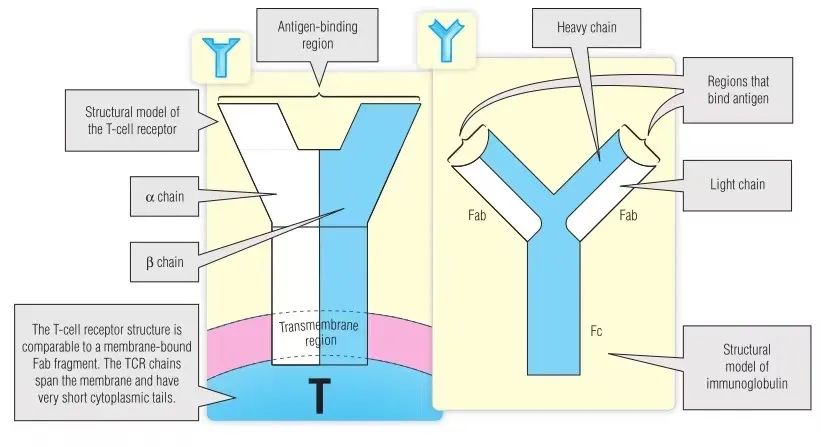
Fig 7.1 T细胞受体(TCR)的结构与免疫球蛋白的比较
T细胞受体的结构
TCR是异二聚体膜蛋白。TCR包括两种类型的受体,分别是αβ链受体和γδ链受体,携带有两种受体的T淋巴细胞的比例分别约占人类总T淋巴细胞的95%和5%。这些受体中的每一个链的分子量都在40,000至60,000之间。每条链的细胞外部分由两个域组成(见Fig 7.1)。整体结构类似于免疫球蛋白(Ig)的抗原结合片段(Fab)的结构。距膜最远的TCR域与Ig可变(V)区域相似,而距膜最近的TCR域与Ig恒定(C)区域相似。抗原结合由αβ或γδ链的V结构域产生的位点进行。已经确定TCR细胞外部分的三维(3-D)结构(Fig 7.2),与Ig具有很大相似性。
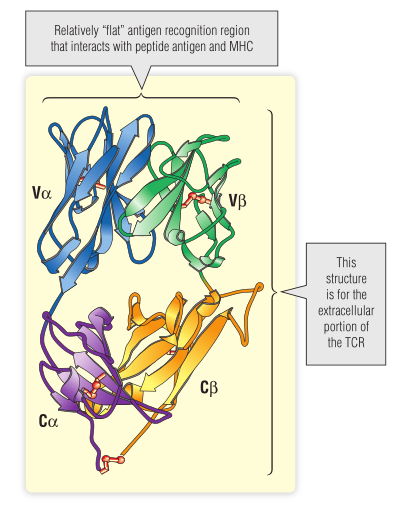
Fig7.2 T细胞受体(TCR)的三维结构
许多具有不同特异性的TCRs已获得蛋白质和核酸序列数据。这些序列的分析结果表明在可变区中存在三个高变(hv)区。3-D结构表明,这些高变区排列为相对平坦的表面(见Fig 7.2),该表面同时与MHC分子和抗原肽的氨基酸残基接触。
αβ T细胞受体
αβ TCR是在具有MHC限制性T细胞上发现的主要TCR(95%)。通常我们提到TCR,指的是αβ TCR。不同类型的辅助T细胞也是αβ T细胞,大多数病毒感染细胞的细胞毒性T淋巴细胞也是如此。αβ TCR识别由MHC分子呈递的肽抗原(第2、3、9和10章)。
γδ T细胞受体
γδ TCR存在于少数(5%)人T细胞上。这些细胞在胸腺中作为与表达αβ TCR分子的细胞不同的细胞谱衍生(第15章)。在某些上皮组织中,γδ T细胞数量更多,这导致了以下假设:它们是第一道防线,并可能引发对上皮边界,如皮肤,经常遇到的微生物的反应。同样,γδ T细胞与αβ T细胞在两个重要方面有所不同。首先,他们既可以识别脂质分子也可以识别肽。第二,他们不能总是识别MHC,不是MHC限制性的。
T细胞受体基因多样性的产生
编码TCR链的基因片段的结构非常类似于Ig重链和轻链基因区段的结构。如Fig 7.3所示,α和γ链的基因与Ig κ和λ轻链的基因类似,因为它们仅使用V和J段。β和δ链的基因类似于Ig重链基因,它们使用V,D和J基因段。与Ig基因的不同之处在于,TCR C区域基因较少。例如,只有一个Cα基因,有两个Cβ基因,但它们在功能上似乎是相同的。这与其中C区包括µ,δ,γ,ε和α类以及λ亚型(λ1至λ4)等的Igs相反。
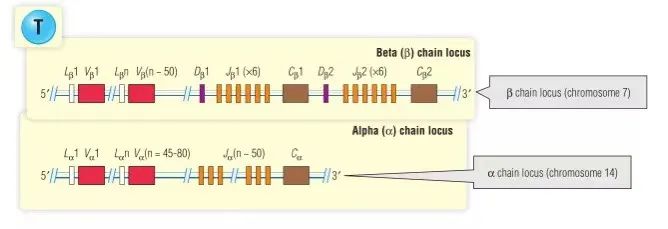
Fig 7.3 人类T细胞受体基因的组织
在抗原刺激T细胞之前产生多样性的机制与B细胞中产生多样性的机制相同(第6章)。但在抗原刺激后,T细胞内途径与B细胞完全不同。抗原刺激后,Ig基因继续多样化(例如通过体细胞超突变和类别转换,将V区连接到另一个C区),而TCR的基因保持不变。
TCR基因在胸腺重排(第15章)。合成TCR链的基本分子步骤与针对Ig轻链和重链所述的分子步骤非常相似(Fig 7.4)。对于B淋巴细胞中的Ig基因,包括RAG-1和RAG-2酶在内的V(D)J重组酶与TCR基因重排有关,这些功能的缺陷会影响B细胞和T细胞(BOX 7.1)。对于α链,V基因区段(如Vα2)和J基因区段(如Jα5)重组以形成V区外显子(如Vα2 Jα5)。V区的转录和Cα外显子一起产生初级RNA转录本。该RNA的剪接产生一个信使RNA(mRNA),该信使RNA在翻译时会产生TCR α链蛋白(参见Fig 7.4),其方式类似于Fig 6.3A和Fig 6.4中针对Ig轻链所述方式。
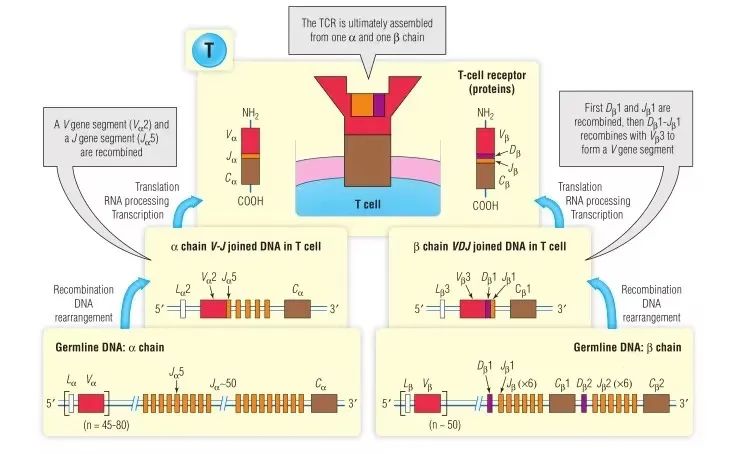
Fig 7.4 人αβT细胞受体的合成与表达
β链的组装与Ig重链的组装相似,首先将αD和αJ基因片段组合在一起,然后将该DJ单元组合到V基因片段上,如Fig 7.4中Dβ1,Jβ1和Vβ3。然后,完整的V区外显子与Cβ1一起转录,形成一级RNA转录物。RNA剪接产生mRNA,翻译后产生TCR的β链(参见Fig 7.4),方式类似于Fig 6.3B和Fig 6.5中针对Ig重链。α和β链像其他与膜结合蛋白一样在粗面内质网中翻译,经过内质网和高尔基体处理后在细胞表面膜上表达。
与免疫球蛋白一样,TCR多样性产生的机制包括:(1)存在多个V区基因,(2)末端脱氧核苷酸转移酶(TdT)的不精确连接和核苷酸添加而产生的连接多样性,(3)随机产生链条组合。与Ig不同,TCR基因中没有体细胞超突变。但是,总的潜在B细胞和T细胞抗原受体种类相似,因为缺乏体细胞超突变被TCR基因中更大的连接多样性的抵消了(第9章)。从理论上讲,TCR数目可能高达1016至1018,其中大部分是由多样性产生。TCR连接多样性的一个例证是TCR α链可用的J片段数量是Ig κ和λ轻链的10倍以上。
抗原的识别
TCR仅出现在细胞表面。T细胞没有像B细胞抗原受体那样表达另一种分泌形式的潜力。Fig 7.5显示TCR如何与自身MHC分子呈递抗原肽相互作用。处理后(第10章)的大多数肽抗原都显示在MHC凹槽(第2、3和8章)中与TCR结合。少数被称为超抗原的物质可独立于抗原加工和呈递而激活T细胞。这些超抗原同时结合MHC II类分子和具有某些特定β链的TCR。通过这样做,它们激活带有该Vβ区域的T细胞,并引起大规模的免疫反应(BOX 7.2)。
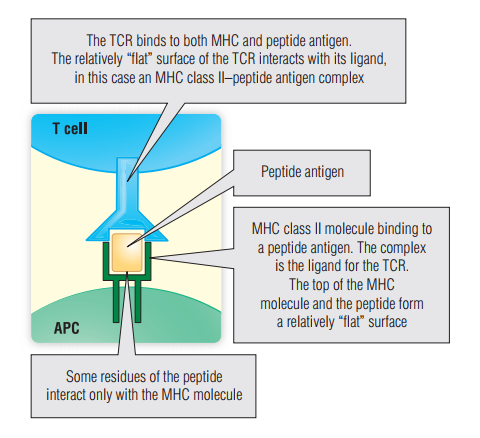
Fig 7.5 T细胞受体(TCR)与肽和主要组织相容性复合体(MHC)的相互作用
对TCR-肽-MHC复合物进行了3-D结构分析。Fig 7.6显示了MHC I类分子(HLA-A2),病毒肽和特异性TCR,该TCR V区的高变序列产生一个“平坦”的表面,该表面与抗原肽的残基以及位于MHC分子α1和α2域中的一些多态氨基酸残基相互作用。
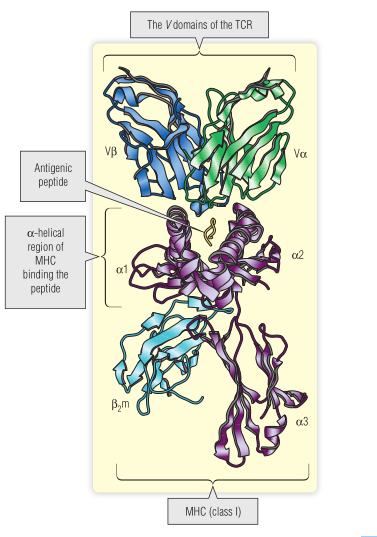
Fig 7.6 MHC-肽-T细胞受体(TCR)复合物的三维结构
涉及T细胞功能的辅助分子
如Fig 7.7所示,几种分子参与T细胞功能。这些分子形成免疫突触的一侧,T细胞利用免疫突触与其他细胞通讯。本章主要关注结合抗原的TCR αβ和γδ链。TCR没有CD3复合物(由四个不同的跨膜蛋白链:γ,δ,ε和ζ)则无法正常发挥受体功能。在TCR结合抗原后,CD3分子通过信号转导至细胞质(第11章)从而使T细胞活化。其他分子,如CD11a等整合素,又称白细胞功能相关抗原1(LFA-1),在T细胞与其靶细胞起黏附作用(见Fig 7.7),而另一些在黏附及信号转导中起作用。其中最重要的是CD4和CD8,它们通过分别与II类或I类MHC分子结合来稳定TCR-肽-MHC复合物,以及通过将酪氨酸激酶(Src家族的成员Lck)带到CD3和ζ蛋白的细胞质尾部附近从而促进信号转导和细胞激活,增强特定T细胞应答。携带CD4的T细胞被称为T辅助细胞,因为它们通常促进其他细胞应答。然而,T辅助细胞的一个子集,称为调节性T细胞(Tregs),能够抑制其他T细胞。携带CD8的T细胞具有杀伤功能,如诱导病毒感染细胞凋亡,也被称为细胞毒性T淋巴细胞(CTL)。
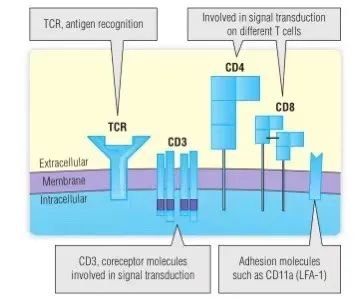
Fig 7.7 参与T细胞功能的辅助分子
免疫突触与神经突触的不同之处在于它是一个瞬时结构。例如,当T细胞识别抗原呈递细胞(APC)上的肽抗原时,突触会形成几个小时。在此期间,两个细胞之间双向通信。一旦T细胞对抗原产生反应,突触就会分解并释放T细胞与其他细胞相互作用,从而对抗原产生反应(Fig 7.8)。这些过程将在第16章中进一步描述。

Fig 7.8 该图显示了CD4+T细胞和抗原提呈细胞之间的免疫突触
BOX 7.1 重组激活基因(RAG-1,RAG-2)产物缺陷,导致常染色体隐性重症联合免疫缺陷
一个三周大的女孩被她的父母带到医院。自出生后婴儿体重减轻了10%,呼吸困难。父母被认为是近亲,没有其他值得注意的东西。经检查,婴儿存在广泛的念珠菌皮肤感染、呼吸衰竭。胸片显示耶氏肺孢子虫肺炎的特征性表现,淋巴细胞计数较低。当使用流式细胞术(第5章)计数各种类型的淋巴细胞时发现T细胞和B细胞完全缺失。有一些自然杀伤(NK)细胞。基因测试确认由RAG1和RAG2基因突变引起的常染色体隐性重症联合免疫缺陷(ARSCID)。婴儿被转诊干细胞移植。
重组激活基因(RAG)酶启动B细胞中的免疫球蛋白基因重排(第6章和第14章)。如本章所述,它们在T细胞中对于T细胞受体基因重排也很重要(第15章)。RAG-1或RAG-2的缺陷导致AR-SCID,这一事实证明其重要性。由RAG1和RAG2突变引起的AR-SCID形式与该疾病的其他形式不同,因为T和B细胞完全缺乏。但在血液循环中发现了NK细胞。因此这种疾病称为TB-SCID。
TB-SCID是一种罕见综合征,如果没有来自人类白细胞抗原(HLA)兼容供体的骨髓移植(BMT),2岁时会致命。该病在最初几周内以淋巴细胞减少和反复感染的形式出现在婴儿中,只能检测到很小的胸腺。这些婴儿反复发作肺炎,中耳炎和皮肤感染,并持续感染机体如白色念珠菌和耶氏假丝酵母菌。对于父母和儿科医生来说孩子处于极大的危险之中。但是,如果足够早地进行BMT,超过80%的儿童可以存活。SCID作为一种综合症很少见,TB-SCID更为罕见。目前BMT之后有数百个人还活着,可以纠正各种形式的SCID。BMT将在第34章中进一步讨论。
BOX 7.2 超抗原和中毒性休克综合症
一名23岁的女性在工作中晕倒后被送往急诊。患者出现大面积红疹,高温(38.9°C)和低血压(收缩压为86毫米汞柱)。据悉,她在月经期间使用了卫生棉条。急查血常规显示她有急性肾功能衰竭和肝炎,对中毒性休克综合症进行了初步诊断。拿去棉条,补液使患者苏醒并恢复健康。在接下来的几天中,血培养为阴性。2周后她的手掌皮肤脱皮。这些特征可以确认中毒性休克综合征的回顾性诊断。
某些细菌和病毒产生的某些蛋白质无需加工即可被表达αβ T细胞受体(TCRs)的T细胞识别。它们直接(但在抗原识别位点之外)与许多TCR Vβ序列结合激活T细胞。它们还与MHC II类分子的外表面结合(Fig 7.9)。由于它们与许多T细胞的TCR Vβ区的结合,超抗原能够激活1%-20%的T细胞,并导致血液中存在高水平的细胞因子。高水平的细胞因子被称为细胞因子风暴,会导致以中毒性休克综合症为特征的低血压和多器官衰竭。请注意局部感染(在这种情况下为受感染的棉条)引起威胁生命的症状。感染性休克是引起细胞因子风暴和血压降低的另一个原因(第21章)。在感染性休克的情况下,感染通常很普遍,血培养通常是阳性。
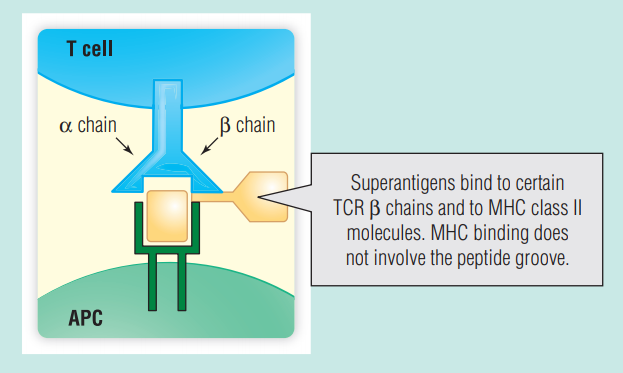
Fig 7.9 MHC II类分子和T细胞受体TCR
BOX 7.3 分化分子簇
在免疫学中,细胞表面分子称为CD,如CD3。该缩写代表分化簇,是发现这些分子时首先使用的术语。到目前为止,已经描述了数百种CD分子。最著名的是CD4,它是HIV附着于T细胞的细胞受体。检测CD4+T细胞的数量可以以评估HIV介导的免疫缺陷疾病的病情进展(第33章)。
— THE END —
▉ 往期精彩内容推荐
GraphPad+AI制作可发表级别的柱状图-OncoLab实验室
本篇文章来源于微信公众号: OncoLab
 微信扫一扫打赏
微信扫一扫打赏
 支付宝扫一扫打赏
支付宝扫一扫打赏