导 读
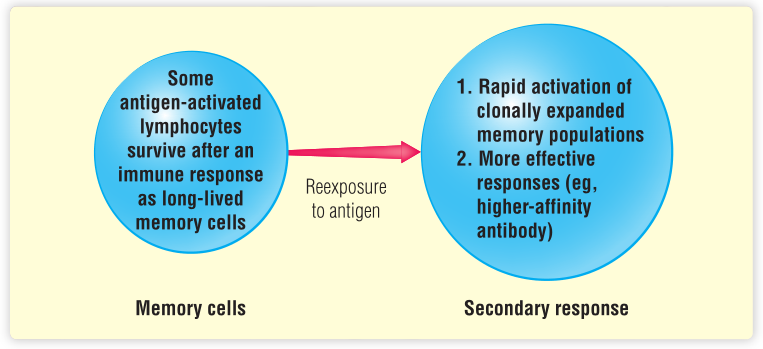
适应性免疫系统的标志之一是“记住”曾经与抗原的接触(第2章)。如上图所示,本章描述了记忆细胞以及记忆性免疫反应是如何产生的,并解释了免疫记忆在再次遇到病原体时体现出的个体优势。免疫记忆在保护机体免受病原体侵袭方面发挥着非常重要的作用,宿主在第二次遇到病原体时可启动二次免疫反应。这种二次免疫反应曾在第2章中讨论过,其具有以下特征:
-
反应更快(几小时即可产生免疫反应);
-
反应更激烈,产生更多抗体和更高亲和力的特异性抗体(Fig.2.7);
-
更具特异性,例如,T细胞中TH1或TH2,免疫球蛋白中的IgG或IgE。
如第2章和第4章(另见第25章)所述,接种疫苗就是利用免疫记忆,针对天花和脊髓灰质炎等传染病的疫苗一直是医学科学最重要的成就之一。随着医疗卫生条件的改善,疫苗极大地提高了世界各地人民的生活质量。虽然关于免疫记忆的利用在医疗上已经取得了巨大的成功,但关于免疫记忆是如何建立和维持的,仍然有很多需要解释的地方。本章总结了长期免疫记忆中最重要的内容。
长期免疫记忆
长期免疫记忆指的是对过去遇到的抗原产生更强和更有效的免疫反应的能力。长期记忆的产生被认为是由于机体内存在通过克隆扩增产生的抗原特异性B和T淋巴细胞,它们会持续多年保持静止状态,甚至会持续终生。例如,50年前接种过预防天花疫苗的个体体内的痘苗病毒特异性T细胞被证明是有效的。关于居住在与世隔绝的岛屿社区的人群的研究表明,由抗体介导的人体对麻疹病毒的抵抗作用持续了65年以上。
B细胞记忆
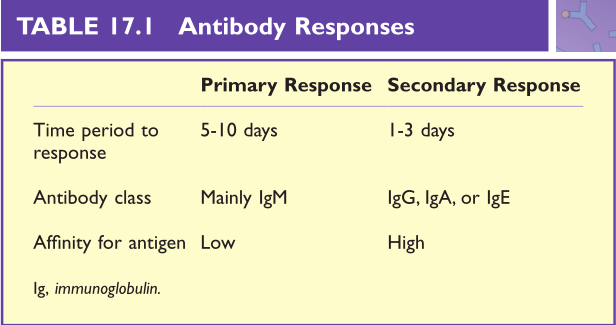
Table17.1 抗体反应
被抗原激活的幼稚B细胞分化为可以分泌抗体的效应B细胞(浆细胞)(第14章)。在初级免疫反应中,浆细胞分泌相对低亲和力的抗体,而在次级免疫反应中分泌亲和力相对较高的抗体。
初次接触抗原后,浆细胞可能会在数月内一直分泌抗体。这对人体是有帮助的,因为这意味着病原体进入人体的时候体内已有抗体存在(Box 17.1)。
体细胞超突变发生在次级免疫应答过程中,可以促使产生与抗原结合亲和力更高的结合位点(第6章和第14章)。免疫球蛋白(Ig)类别转换可以在初次免疫应答期间发生,但通常初次应答主要是IgM,而再次应答可以是IgG、IgA或IgE(Table 17.1)。
随着时间的推移,抗体水平下降,记忆B细胞需要在再次接触病原体时分泌新的抗体。记忆B细胞很可能来源于激活的B细胞,其可变区基因片段DNA发生了可遗传变异(体细胞超突变),从而产生更有效的抗体(Fig 17.1)。
记忆B细胞保留了高亲和力的抗原受体,这种受体是通过连续的体细胞遗传变化而产生的。随后暴露于抗原时,做出反应的是该记忆细胞而不是别的幼稚B细胞。证明这一点的证据来自于观察,在早期接触过病原体(如麻疹病毒)的人再次接触病原体时,会对病原体上的相同表位做出反应,即使新的表位具有抗原性,他们也不会对其做出反应。这一观察结果与第二次及随后的抗原暴露结果一致,即激活了先前存在的记忆细胞,而不是诱导新的幼稚细胞分化。
T细胞记忆
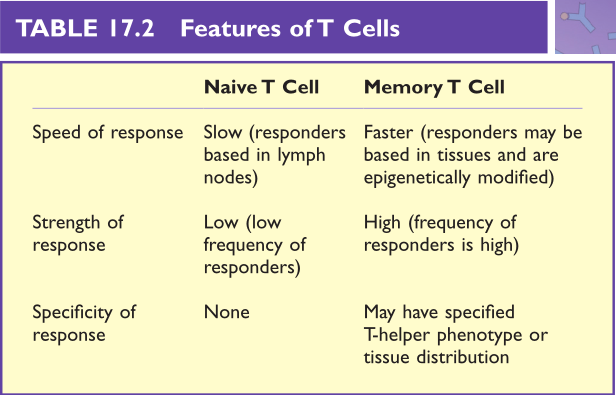
Table 17.2 初始T细胞与记忆T细胞的特征比较
记忆T细胞是在应对感染的初次免疫反应过程中产生的。目前尚不清楚它们是在成为效应性T细胞之前由激活的T细胞产生的,还是由效应性T细胞产生的,或者两者兼而有之。在获得记忆表型的过程中,T细胞经历了具有重要作用的表观遗传修饰。在增殖过程中,这些基因表达模式被传递给子代细胞。第16章解释了这些表观遗传修饰是如何获得的。
T细胞记忆的基本特征是对感染的反应更快、更激烈、更具特异性(Table 17.2)。出于以下几个原因,记忆性T细胞比幼稚T细胞对机体感染做出的反应更快。首先,幼稚T细胞表达L-选择素使得它们可以循环到淋巴结(第13章)而不是其他组织。由此幼稚T细胞处于与抗原提呈细胞(APC)(如树突状细胞)相互作用的最佳位置。相反,记忆T细胞与效应T细胞非常相似,它表达一系列细胞粘附分子,使它们能够进入各种组织,当感染发生时记忆T细胞可迅速到达感染部位。此外,表观遗传的变化意味着基因的转录速度更快。例如,细胞因子基因可能更松散地排列在组蛋白周围,使它们更容易发生转录。综上所述,记忆T细胞在组织中经过表观遗传修饰后,对感染的应答更快。
记忆T细胞反应比初级T细胞反应更激烈,因为在记忆T细胞群中有更多的T细胞带有抗原特异性受体。T细胞受体基因在胸腺中随机重排。经过阳性、阴性选择后,T细胞识别特定抗原肽的几率非常低(大约在1:10,000到1:100,000之间)。在初级T细胞反应中,特异性T细胞增殖。在初始应答的高峰期,特异性T细胞的数量可能会增加10000个,但这些细胞大部分会发生凋亡(Fig 17.1),只有小部分细胞存活(1:100~1:1000)并产生长期免疫记忆。
记忆性T细胞反应比初级免疫反应更特异。这主要适用于CD4+T辅助细胞(TH),当它们成为记忆细胞时,将会获得TH1,TH2,TH17,以及其他可能的表型。这些特殊的表型,加上受限的循环模式,形成了非常特殊的反应模式。例如,流感特异性的TH1记忆细胞往往会在肺中富集,恰好是在需要它们的地方。
上一章中,非胸腺依赖的抗原B细胞的免疫记忆能力到婴儿后期才会成熟。出于这个原因,个体生命的最初两年感染率最高。从2岁到20岁左右,T细胞和B细胞的记忆才逐步建立起来,此时个人被感染率处于中等水平。从20岁左右开始,受感染率降低。T细胞的记忆可以持续至少50年,这可能是通过T细胞克隆及其子代细胞的存活,而不是单个记忆细胞的长期存活。(见第25章中接种天花疫苗后的免疫)。不幸的是,大约60年后,随着免疫系统的衰老(第33章),感染率会再次增加。
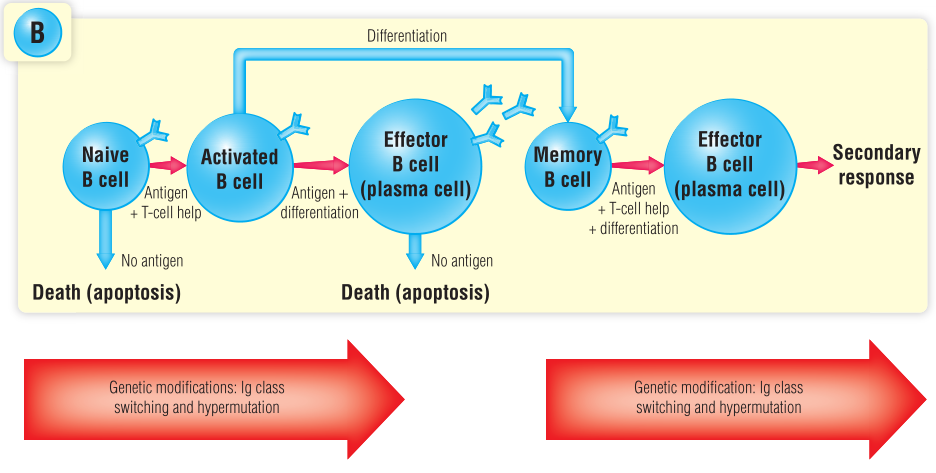
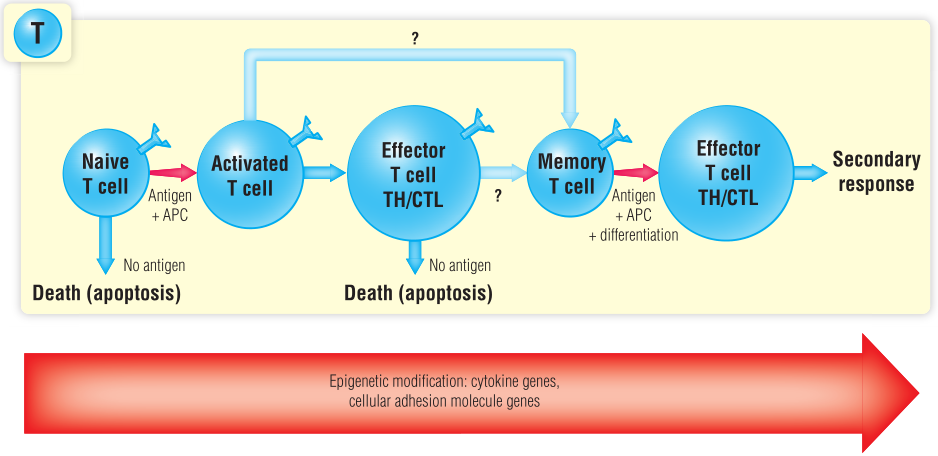
Fig 17.1 记忆细胞的分化途径
淋巴细胞动态平衡
鉴于我们有可能对我们遇到的每一次外来抗原接触保持免疫记忆,即使在每一次免疫反应中产生的记忆细胞数量很少(通常是大量的),最终也会出现一个问题,即是否有合适的身体部位来维持这些细胞。然而,血液和淋巴组织能容纳的细胞数量有限。但幸运的是,像其他淋巴细胞一样,大多数效应细胞和记忆细胞最终似乎都会死亡,可能是通过程序性细胞死亡(凋亡)来诱导死亡。涉及细胞凋亡的机制将在第22章中进一步详细描述。通过细胞死亡可以使初级免疫反应中淋巴细胞的大量克隆性扩增与在有限数量淋巴细胞的情况下维持记忆细胞之间保持平衡(动态平衡)。
大多数被激活的效应细胞随着抗原的消除而死亡,同时共刺激分子和T细胞生长因子白细胞介素2(IL-2)的水平下降。如果没有如此,则促凋亡蛋白的水平就会增加(Box 17.2),促使细胞凋亡。
抗原特异性记忆细胞的克隆体可以存活更长时间。这些记忆细胞需要争夺空间和可用的生长因子(如细胞因子)来帮助它们存活下来。鉴于这些细胞资源的供应有限,来自于某些抗原刺激产生的记忆细胞可能不会长期存活,存活下来的记忆细胞更适合在这些条件下生存。例如,他们对细胞因子和生长因子的需求减少;因此,他们在生存能力方面更具竞争力。这可能意味着对某些抗原的免疫记忆可能比对其他抗原的记忆更稳定。
细胞凋亡
细胞凋亡是通过受体-配体相互作用,主要是Fas-FasL相互作用触发的(Fig 17.2)。
FasL(例如在杀伤T细胞上表达)与Fas(例如在靶细胞上表达)的结合,在靶细胞中触发一系列的细胞内生化反应。Fas与“死亡受体途径”中的几种蛋白质相互作用,最终激活一种被称为caspase的蛋白水解酶,该酶在激活caspase级联中其他的几种蛋白水解酶方面起着关键作用。这种蛋白水解级联在某种程度上类似于细胞激活的激酶级联(第11章)。Caspase级联的关键步骤是激活细胞质酶,caspase可激活DNA酶(CAD),然后它可以迁移到细胞核,将DNA裂解成小片段,引起细胞凋亡(见Table 17.3和第22章)。
已经发现了几个促进细胞死亡的基因,以及几个抑制细胞死亡的基因(Box 17.2)。死亡抑制基因家族包括BCL基因【最初在B细胞淋巴瘤中发现(第35章)】,其是典型的死亡抑制或抗凋亡基因。对长期记忆细胞存活的其中一种解释是,它们的BCL蛋白家族的表达水平高于正常水平。这一特性保护它们免受抗原诱导的细胞死亡,并促使它们的长期生存。
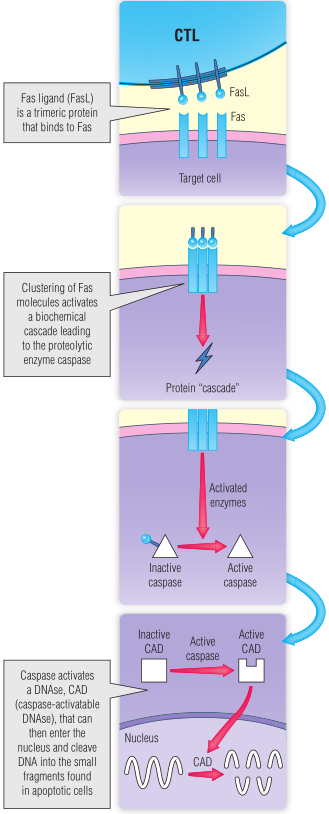
Fig 17.2 细胞死亡途径
BOX 17.1医学院入场券:免疫记忆
安妮和贝琳达都被医学院录取了。为了能够上学,他们的乙肝抗体水平必须超过100mIU/mL(不同国家的规定有所不同)。安妮在6周前完成了最初的三针疫苗接种(见Fig 2.7)。血液检测显示她的抗体水平是135Miu/mL,所以她可以开始上学了。
贝琳达在4年前夏天做实验室助理时接种了乙肝疫苗。当时她进行了血液检测,结果显示抗体水平为180mIU/mL。然而,当她在医学院筛查时再次接受检测时,她的抗体水平已降至43mIU/mL。她接种了加强疫苗,两周后她的水平已经上升到380mIU/mL,所以她被允许开始上学。
为了保护肝脏,血液中的抗体需要处于高水平(超过100mIU/mL)。尽管抗体水平可能会随着时间的推移而下降,但在大多数人中,三次乙肝疫苗注射的疗程后可以达到要求的标准。增强疫苗诱导记忆B细胞成熟为浆细胞,并再次开始分泌抗体。因为这是一种记忆反应,所以抗体水平更高,比初次反应更快。
BOX 17.2凋亡:程序性细胞死亡
细胞凋亡是清除多余或受损细胞的一种机制。这是一个由濒临死亡的细胞启动的进化保守的过程,它代表了一种受控的细胞破坏形式。目前已经确定了几个促进或抑制细胞凋亡的基因(Table17-3)。抗凋亡基因有一些特征,如比其他基因存活时间更长。Bcl-2是重要的抗凋亡基因。Bcl-2型的抗凋亡基因似乎通过提高细胞的凋亡阈值来发挥作用;即表达Bcl-2蛋白的细胞所需的凋亡启动剂剂量大于没有Bcl-2蛋白的细胞所需的凋亡启动剂剂量。
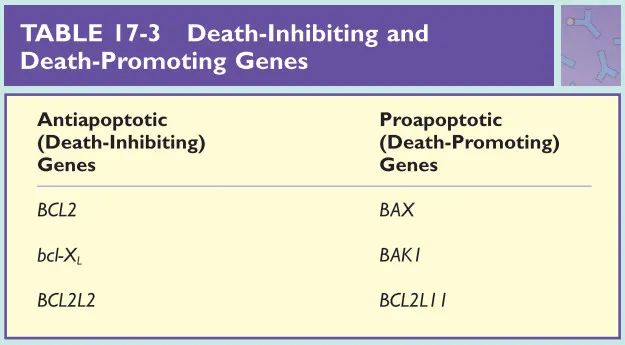
Bcl-2基因最初是作为一种哺乳动物癌基因被检测到,它与人类染色体14和18之间的易位有关,在超过70%的淋巴瘤中可以检测到这种基因。染色体易位导致Bcl-2的表达增加,这导致由该染色体易位产生的B肿瘤细胞具有相对高的抗凋亡能力(第35章)。
— THE END —
▉ 往期精彩内容推荐
基础免疫学系列
肿瘤免疫学系列
即将开始,敬请期待!
生信入门学习笔记系列
基础实验技术系列
Annexin V/PI染色检测细胞凋亡-OncoLab实验室
仪器操作系列
蔡司LSM800激光共聚焦显微镜操作视频-中西医结合基础研究中心
IN Cell Analyzer2200高内涵细胞成像分析系统操作教程-中西医结合基础研究中心
文献分享系列
年度巨献:神刊CA发布最新癌症数据:全球1000万人死于癌症,中国占比超3成
HLA-I分子加工和递呈抗原功能受损是免疫检查点抑制剂耐药的重要原因
作图投稿选刊系列
GraphPad+AI制作可发表级别的柱状图-OncoLab实验室
PubMed中如何按影响因子筛选检索结果并显示影响因子及分区?
如何使用EndNote软件插入参考文献(附6813种杂志参考文献格式文件)

长按上方二维码即可关注本号~
本篇文章来源于微信公众号: OncoLab
 微信扫一扫打赏
微信扫一扫打赏
 支付宝扫一扫打赏
支付宝扫一扫打赏